
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Расщепление нуклеопротеинов в пищеварительном тракте
Расщепляются на белок и нуклеиновые кислоты (НК) Расщепление белков – протеазы. НК – нуклеазы (фосфодиэстеразы) панкреатического сока – гидролизуют 3’, 5’-фосфоэфирные связи между нуклеотидами. Под действием РНКаз образуются моно-, ди-, три- и олигонуклеотиды. Под действием ДНКаз – в основном ди- и олигонуклеотиды, реже мононуклеотиды. Мононуклеотиды в кишечнике расщепляются под действием фосфатаз (кислая, щелочная) с образованием нуклеозидов и фосфата (разрушается фосфоэфирная связь). Частично распадаются в клетках слизистой оболочки кишечника под действием нуклеотидаз с образованием нуклеодизов и фосфата. Образованные нуклеозиды через промежуточные продукты ресинтезируюся (90% нуклеозидов в организме – за счет ресинтеза, 10% – синтез de novo). Дальнейшие превращения нуклеозидов: · всасываются в тонком кишечнике и могут быть использованы для синтеза НК организма · или дальше расщепляются фосфорилитически (не гидролитически! ) Е: нуклеозидфосфорилазы с образованием азотического основания и рибозы-1-фосфата.
Внутриклеточнное расщепление НК – под действием нуклеаз (до нуклеозидов, далее до азотистых оснований и рибозо-1-Р). Далее азотистые основания могут быть катаболизированы, но большая часть – на ресинтез нуклеотидов, т.к. синтез – очень энергозатратный процесс. Классификация нуклеаз
II. В зависимости от субстратной специфичности: · Рибонуклеазы (РНКазы) · ДНКазы · Неспецифические нуклеазы III. В зависимости от места гидролитического расщепления фосфодиэфирной связи: · А-тип: расщепление связи между фосфатом и 5’-атомом углерода остатка рибозы или дезоксирибозы (с образованием нуклеозид-3’-Р) · В-тип: расщепление между фосфатом и 3’-атомом углерода рибозы или дезоксирибозы (с образованием нуклеозид-5’-Р) IV. По типу расщепляемой цепи: · Расщепляющие одноцепочечные НК · Расщепляющие двуцепочечные НК V. В отдельный тип выделяют рестриктазы – нуклеазы, узнающие определенные последовательности нуклеотидов: · Катализируют гомологичный разрыв («тупые» концы) · Катализируют гетерологичный разрыв («липкие» концы)
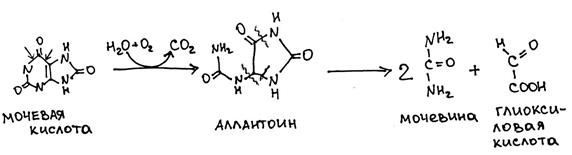
Катаболизм пуриновых оснований (А и Г) Продукты: 1. Мочевая кислота – у млекопитающих, птиц, некоторых рептилий 2. Аллантоин – у рептилий, некоторых млекопитающих 3. Аллантоиновая кислота и мочевина – у рыб
*Окисление гипоксантина и ксантина осуществляется ксантиноксидазой с образованием активных форм кислорода (H2O2). У ряда животных (но не у человека! ) и большинства растений может осуществляться дальнейшее окисление до аллантоина и мочевины:
Катаболизм пиримидиновых оснований (Ц и Т) Пиримидиновые основания подвергаются НАДФН-зависимому восстановлению до NH3 и β -аминокислот. Продукты: CO2, NH3, мочевина, β -аланин (далее используется для синтеза ансерина, карнозина, образования КоА), β -аминомасляная кислота. Репликация ДНК. Основные типы ДНК-полимераз, их структура, ферментативная активность, роль во внутриклеточных процессах. Контроль точности воспроизведения ДНК.
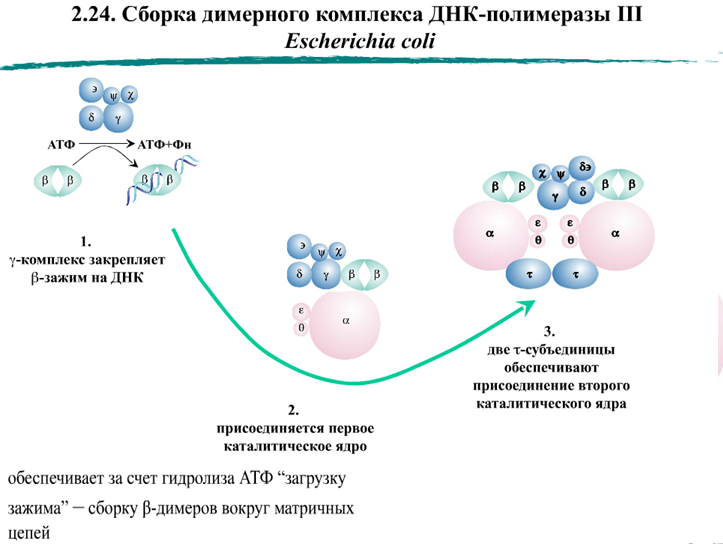
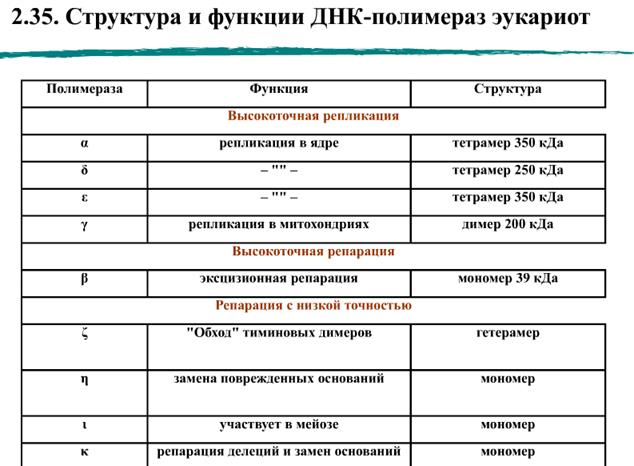
Бактерии E. coli имеют пять ДНК-полимераз:
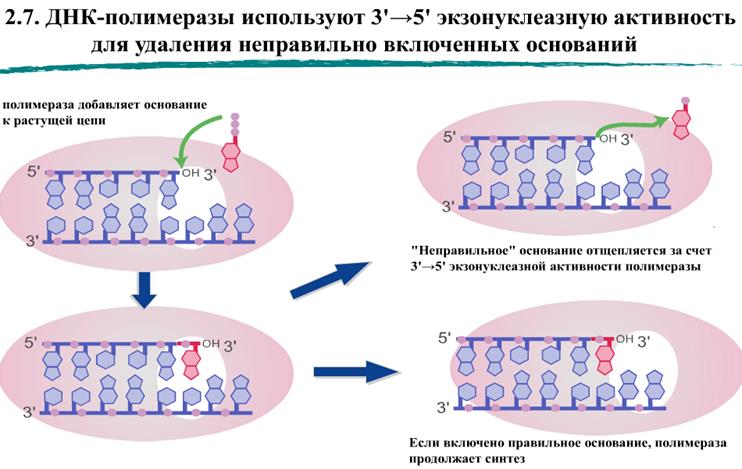
Кроме полимеразной, ДНК-полимеразы могут иметь экзонуклеазные активности для удаления нуклеотидов с концов цепи ДНК. Экзонуклеазные активности ДНК-полимераз на несколько порядков повышают точность синтеза ДНК
Репарация ДНК. Механизмы эксцизионной репарации ДНК (эксцизия нуклеотидов, оснований). Пострепликативная коррекция неспаренных оснований. Репарация генетических повреждений — свойство живых организмов восстанавливать повреждения, возникшие в ДНК в результате воздействия разнообразных мутагенных факторов как радиационной, так и химической природы. Оно было открыто относительно недавно, менее полувека назад. Эксцизионная репарация. Существуют более сложные реакции восстановления, напоминающие хирургические вмешательства в структуру ДНК, когда поврежденные участки вырезаются из цепи ДНК (отсюда происходит и термин “эксцизионная репарация”, от англ. excision — вырезание), а затем образовавшиеся бреши заполняются неповрежденным материалом.
К настоящему времени описано много типов ферментов-гликозилаз, каждый из которых узнает разнообразные поврежденные основания (такие, как метилированные, окисленные, восстановленные, дезаминированные основания, основания, связанные с группировками, и т.п.. Гликозилазы присоединяются к ним, рвут гликозидные связи между модифицированным основанием и сахаром дезоксирибозой, за счет чего образуются АП-сайты. АП- сайт распознается теперь другим ферментом, АП- эндонуклеазой. Последние найдены у бактерий, растений, животных, включая человека. Как только в нити ДНК возникает разрыв, в работу вступает еще один фермент — фосфодиэстераза: он отщепляет от ДНК ту сахарофосфатную группу, к которой теперь не присоединено основание. Появляется брешь в одной цепи ДНК размером в один нуклеотид. Напротив бреши в противоположной нити ДНК расположен не поврежденный нуклеотид, и следующий фермент — ДНК полимераза I вставляет в брешь комплементарный ему нуклеотид, присоединяя его к свободному З’ОН-концу. Чтобы соединить два свободных конца (З’ОН-конец вставленного нуклеотида и 5'-конец, ранее образовавшийся при разрыве нити ДНК АП-эндонуклеазой), вступает в действие еще один фермент — полинуклеотидлигаза. Теперь вся структура ДНК полностью восстановлена: неправильное основание удалено, сахарофосфат, к которому это основание было прикреплено, вырезан из нити ДНК, брешь заполнена правильным нуклеотидом, и все однонитевые разрывы залечены. Поскольку последовательность реакций запущена в действие путем расщепления гликозидной связи, этот вид репарации получил название “вырезание оснований с помощью гликозилаз”.
Другим типом эксцизионной репарации является более сложная и энергетически более дорогая реакция вырезания не просто поврежденного основания, а значительного участка цепи ДНК перед и позади повреждения (рис. 1). Эту реакцию в клетках Е. coli выполняет мультиферментный комплекс, содержащий эндонуклеазы, кодируемые тремя генами: uvrA, uvrB и uvrC (названия генов даны по первым буквам слов ultra violet repair). Комплекс получил название “эксинуклеаза”. Процесс был обнаружен в клетках человека в лаборатории Р. Сетлоу в США и в моей группе (В. Сойфер, Н. Яковлева-Сойфер и А. Мустафина) в 1970 году. Поиск эксцизионной репарации у растений оказался более трудным. Сначала американцы Дж. Троско и Э. Мансур не смогли ее обнаружить и в 1969—1973 годах заявили, что высшие растения либо не получили в ходе эволюции этот важный механизм, либо в ходе той же эволюции утеряли его. Однако в 1973—1978 годах в моей лаборатории, когда я еще работал в Москве, эксцизионная репарация у растений была найдена, а затем изучены процессы повреждения ДНК растений облучением и алкилированием. Было показано, что подавление репарации ведет к резкому увеличению числа мутаций хромосом, замедлению роста и развития растений и к другим нежелательным последствиям. В последние годы эксцизионную репарацию интенсивно исследовали ученые многих стран мира. В частности, было показано, что человеку требуется в среднем в четыре раза больше ферментов репарации, чем бактериям (эксинуклеаза состоит по крайней мере из 17 белков, застройка бреши идет с участием ДНК полимераз а или £ ), а вырезаемый из поврежденной ДНК кусок имеет длину не 12, а 29 нуклеотидов и т.д. |
Последнее изменение этой страницы: 2017-04-13; Просмотров: 1138; Нарушение авторского права страницы
 Цитидин + H3PO4 цитизин + рибоза-1-фосфат
Цитидин + H3PO4 цитизин + рибоза-1-фосфат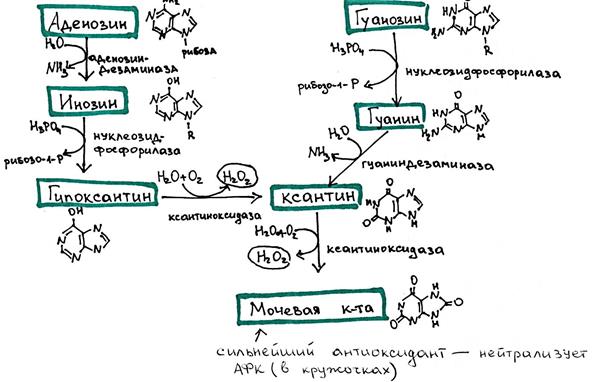

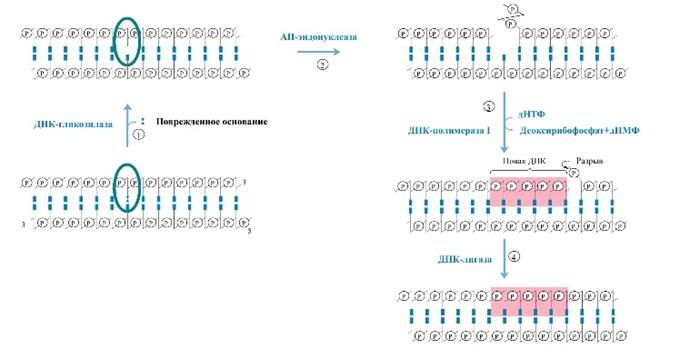 А. Вырезание поврежденных оснований гликозилазами и застройка АП-сайтов.
А. Вырезание поврежденных оснований гликозилазами и застройка АП-сайтов. 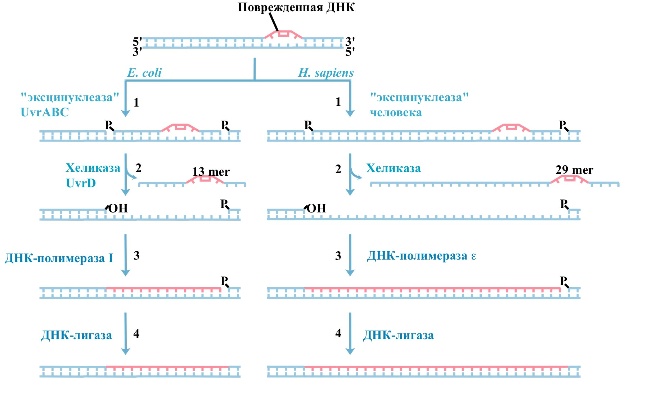 Б. Вырезание нуклеотидов.
Б. Вырезание нуклеотидов.