|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Транскрипция. Матричная РНК, её структура и функциональные участки, различия у про- и эукариот. РНК-полимеразы про- и эукариот.
Транскрипция – синтез цепи РНК на матрице одной из цепей ДНК-дуплекса. Эта РНК идентична кодирующей ДНК (кДНК) и комплементарна матричной ДНК цепи. Транскрипция осуществляется ДНК-зависимыми РНК полимеразами (РНК-пол). Активности РНК-пол: · Расщепление и восстановление двуцепочечных ДНК · Удержание в правильной ориентации расплетенных цепей ДНК и РНК-продукта · Катализ добавления нуклеотидов к растущей РНК · Отщепление 3’-нуклеотидов в случае ошибки и возобновления синтеза (при участии дополнительных факторов) Этапы: 1. Распознавание матрицы начинается со связывания РНК-пол с двуцепочечным участком ДНК в области промотора (связывание TBP (ТАТА-связывающий белок) с ТАТА-боксом). Образуется «закрытый» комплекс, который под действием факторов инициации превращается в «открытый». Факторы инициации вызывают изменение конформации РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка, в которой матрица доступна для инициации синтеза цепи РНК. 2. Инициация. Происходит синтез первых нуклеотидных звеньев. Может затянуться из-за абортивной инициации (высвобождение коротких транскриптов; синтез начинается заново, пока полимераза не сдвинется на 8-10 нуклеотидов). После того как синтезирован фрагмент из 8-10 нуклеотидных остатков, σ -субъединица отделяется от РНК-полимеразы (в 70% случаев, т.к. для инициации не нужна), а вместо неё к молекуле фермента присоединяются несколько факторов элонгации. 3. Элонгация. Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК. 4. Терминация. Распознавание специфической последовательности ДНК (терминатор), прекращающей дальнейший рост цепи РНК. Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК), комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
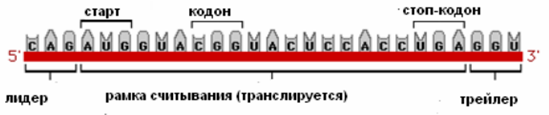
I. Структура мРНК у про- и эукариот мРНК (и РНК) – промежуточная копия последовательности ДНК, несущей информацию о строении белка. Прокариоты Размеры синтезированной мРНК у бактерий практически совпадают с размерами гена – средний размер бактериальной мРНК – 1 500 нуклеотидов. Если транскрибируется оперон (совокупность генов), то длина мРНК может достигать 5‐ 15 т.н. Бактериальные мРНК очень нестабильны (время полураспада около 2 мин). Не претерпевают существенных модификаций. Трансляция и транскрипция не разобщены во времени – как только синтезируется свободный 5’-конец определенной длины, рибосомы садятся на него и начинают трансляцию. Деградация осуществляется в 2 ступени: · Серия эндонуклеотических разрезов (в направлении 5’-3’) · Экзонуклеазная деградация небольших фрагментов (3’-5’). Структура:
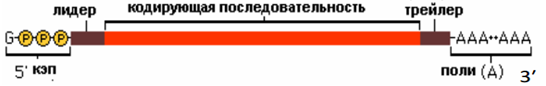
1. Лидер – 5’ не транслируемый участок (5’UTR) 2. Трейлер – 3’ не транслируемый участок (3’UTR) 3. Рамка считывания – участок мРНК, кодирующий синтез полипептида (от старт-кодона до стоп-кодона). Эукариоты У эукариот длина мРНК значительно укорачивается после созревания (сплайсинга), в ходе которого удаляются интроны. Длина типичной зрелой мРНК – от сотен до нескольких тыс нуклеотидов. Относительно стабильна (время полураспада у животных 4-24 ч). Транскрипция и трансляция разобщены во времени и пространстве. Транскрипция и процессинг осуществляется в ядре; трансляция – в цитоплазме. Процессинг: 1. Модификация концов (защищает от экзонуклеаз): · 5’-конец – кэпирование. Фосфатаза фосфорилирует 5’-конец (за счет фосфата ГТФ). Гуанилтрансфераза добавляет G через 5’-5’-связь Метилтрансфераза добавляет от 1 до 3 метильных групп к основанию или рибозе нового концевого гуанидина Функции кэпа: А) для узнавания мРНК при инициации трансляции Б) для защиты 5’-конца от рибонуклеаз, специфически разрезающих фосфодиэфирные связи в направлении 5’-3’ и атакующих незащищенный 5’-конец В) участвует в созревании 3’-конца мРНК Г) осуществляет экспорт мРНК из ядра в цитоплазму Д) участвует в сплайсинге Е) участвует в регуляции транскрипции · 3’-конец – полиаденилирование (добавление к транскрипту поли-А-хвоста из ~200 нуклеотидов). PABP (поли-А-связывающий белок) – определяет длину хвоста, наматывая на себя поли-А-хвост. Специальная эндонуклеаза узнает AAUAAA-участок и на расстоянии ~20 нуклеотидов от него в направлении к 3’-концу делает надрез на пре-мРНК. Затем происходит добавление поли(А)-хвоста – 30-300 адениловых нуклеотидов (безматричный синтез). Функции полиаденилирования: А) способствует экспорту зрелых мРНК из ядра Б) защищает мРНК от действия нуклеаз в цитоплазме, тем самым увеличивая период полураспада (чем длиннее хвост, тем дольше «живет») В) служит в качестве сигнала узнавания для рибосомы 2. Созревание пре-мРНК (сплайсинг) – вырезание интронов (по типу лассо) и сшивание экзонов в том же порядке. 3. Альтернативный сплайсинг – форма сплайсинга, обеспечивающая кодирование одним геном структурно и функционально различающихся полипептидов. Происходит соединение экзонов в различной последовательности. У человека 94% генов подвергает альтернативному сплайсингу. 1 ген кодирует множество форм белка. 4. Редактирование РНК – процесс, в ходе которого информация, содержащаяся в молекуле РНК, изменяется путем химической модификации оснований. В настоящее время установлено редактирование тРНК, рРНК и мРНК эукариот. Редактирование РНК в клетках прокариот не описано. · Модификация нуклеотидов (дезаминирование С в U и А в инозин) · Вставка нуклеотидов без матрицы Биологические последствия редактирования мРНК: А) образование пригодной для трансляции мРНК из транскриптов, которые не могут дать функциональный белок Б) редактирование может дать новый белок, отличный по функциональной активности (аполипопротеин В100 – в кишечник САА в UAA и В48 – в печени без редактирования) Структура:
5’-кэп – 7-метилгуанозин; Лидер (5’UTR); Кодирующая последовательность; Трейлер (3’UTR); 3’-поли(А)-хвост.
|
Последнее изменение этой страницы: 2017-04-13; Просмотров: 2320; Нарушение авторского права страницы