|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Химический состав желудочного сока. Патологические компоненты желудочного сока.Стр 1 из 3Следующая ⇒
В-окисление жирных кислот. окисление жирных кислот протекает в печени, почках, скелетных и сердечной мышцах, в жировой ткани. Первая стадия β -окисления — дегидрирование активированной жирной кислоты (ацил-КоА) с образованием β -ненасыщенной жирной кислоты с двойной связью в транс-конфигурации (реакция 1: дегидрирование). При этом оба атома водорода с электронами переносятся от фермента наэлектронпереносящий флавопротеин (ETF). ETF-дегидрогеназа переносит восстановительные эквиваленты на убихинон (кофермент Q), который является составной частью дыхательной цепи. Вторая стадия деградации жирной кислоты состоит в присоединении молекулы воды к двойной связи ненасыщенной жирной кислоты (реакция [2]: гидратирование). На третьей стадии происходит окисление гидроксильной группы при С-3 в карбонильную группу (реакция [3]: дегидрирование). Акцептором для восстановительных эквивалентов является НАД+ который передает их в дыхательную цепь. На четвертой стадии активированная β -кетокислота расщепляется ацилтрансферазой (β -кетотиолазой) в присутствии кофермента А (реакция [4]: тиолитическое расщепление). Продуктами реакции являются ацетил-КоА и активированная жирная кислота, углеродная цепь которой короче на два углеродных атома по сравнению с длиной цепи исходной жирной кислоты. 8.Холестерин, биологическое значение. Биосинтез холестерина до меваловои кислоты. Холестери́ н - органическое соединение, природный жирный (липофильный) спирт, содержащийся в клеточных мембранах всех живых организмов, за исключением безъядерных (прокариоты). Нерастворим в воде, растворим в жирах и органических растворителях. Около 80% холестерина вырабатывается самим организмом (печенью, кишечником, почками, надпочечниками, половыми железами), остальные 20% поступают с пищей. 80% холестерина в организме свободные, а 20% — связанные. Холестерин обеспечивает стабильность клеточных мембран в широком интервале температур. Он необходим для выработки витамина D, выработки надпочечниками различных стероидных гормонов, включая кортизол, альдостерон, женских половых гормонов эстрогенов и прогестерона, мужского полового гормонатестостерона, играет важную роль в деятельности нервной и иммунной системы. Биосинтез:
Патологии липидного обмена. Нормальное содержание общих липидов в сыворотке крови составляет 4-8 г/л. Увеличение общих липидов называется гиперлипемией. 10.Эикозаноиды. Пути образования и биологическая роль в организме. Эйкозаноиды большая группа медиаторов, обладающих широким спектром биологической активности.Предшественником эйкозаноидов является арахидоновая кислота. Депонироваться эйкозаноиды не могут, разрушаются в течение нескольких секунд, поэтому клетка должна синтезировать их постоянно из поступающих в неё соответствующих жирных кислот.Выделяют три основные группы эйкозаноидов: простогландины, тромбоксаны, леикотриены. Пути образования из арахидоновои кислоты: Роль эикозаноидов: Эйкозаноиды участвуют во многих процессах: регулируют тонус ГМК и вследствие этого влияют на АД, состояние бронхов, кишечника, матки. Эйкозаноиды регулируют секрецию воды и натрия почками, влияют на образование тромбов. Разные типы эйкозаноидов участвуют в развитии воспалительного процесса, происходящего после повреждения тканей или инфекции. Такие признаки воспаления, как боль, отёк, лихорадка, в значительной мере обусловлены действием эйкозаноидов. Избыточная секреция эйкозаноидов приводит к ряду заболеваний, например бронхиальной астме и аллергическим реакциям. 11.Биологическая ценность белков, потребность в белке и аминокислотах. Азотистыи баланс и его виды. Биологическая ценность белков. Определяется следующими критериями: 1.Чем ближе аминокислотный состав пищевого белка к аминокислотному составу беков тела, тем выше его биологическая ценность.2.В составе белка должны быть все незамеимые аминокислоты: вал, илей, лей, лиз, мет, тре, три, фен. Исключение какой-либо незаменимой аминокислоты из пищи сопровождается развитием отрицательного азотистого баланса.3.В составе белка установлены пропорциональные величины незаменимых аминокислот, относительно триптофана, принятого за единицу, лиз-5, лей-4, вал-3, 5.Недостаток в пище одной незаменимой аминокислоты ведет к неполному усвоению других аминокислот.Для человека наиболее ценны белки молока, яиц, мяса, т.к. их аминокислотный состав ближе к аминокислотному составу органов и тканей человека.Потребность в белках зависит от пола, веса, интенсивности труда и других факторов. Все эти факторы влияют на суточную потребность в белке(1-1.5г на кг веса) – общее количество белковой пищи, которую человек должен получить в течение светлого времени суток. Азотистый баланс – разница между введением с пищей азота и выведением его в виде конечных продуктов азотистого обмена.Различают 3 вида азотистого баланса: Положительный – количество выводимого из организма азота меньше количества азота, вводимого с пищей. Азот остается в организме и расходуется на синтез белков. Характерен для растушего организма и во время беременности.Отрицательный – количество выделяемого азота превышает количество азота, поступающего в течение суток. Встречается при голодании, белковой недостаточности, тяжелых заболеваниях, когда происходит интенсивный распад белков тела. Отрицательный азотистый баланс наблюдается у пожилых людей. Азотистое равновесие – количество азота выводимого из организма, равно количеству получаемого с пищей азота. Характерно для здорового взрослого человека.
12.Переваривание и всасывание белков в ЖКТ. В желудке имеются все условия для переваривания белков. Во-первых, в желудочном соке содержится активный фермент пепсин. Во-вторых, благодаря наличию в желудочном соке свободной соляной кислоты для действия пепсина создается оптимальная pH среды. Чистый желудочный сок имеет кислую среду (pH 0, 9-1, 6), а оптимум pH действия пепсина равен 1, 5-2, 5. Следует особо указать на существенную роль соляной кислоты в переваривании белков: она переводит неактивный пепсиноген в активный пепсин, создает оптимальную среду для действия пепсина, в присутствии свободной НС1 происходят набухание белков (увеличение поверхности соприкосновения фермента с субстратом), частичная денатурация и, возможно, гидролиз пищевых белков. Пепсин гидролизует преимущественно пептидные связи, образованные аминогруппами ароматических аминокислот (фенилаланин, тирозин). Он расщепляет практически все природные белки. Исключение составляют некоторые кератины, протамины, гистоны и мукопротеиды. Дальнейшее превращение белков пищи осуществляется в тонком кишечнике, где на белки действуют ферменты панкреатического и кишечного соков. В поджелудочной железе вырабатываются три белковых фермента: трипсин, химотрипсин и карбоксипептидаза. Первые два фермента действуют на белки аналогично пепсину, но в отличие от него они разрывают другие внутренние пептидные связи. Кроме того, они активны в слабощелочной среде (оптимум pH действия их составляет 7, 2-7, 8). Благодаря гидролитическому действию всех трех эндопептидаз (пепсина, трипсина, химотрипсина) на белки образуются различной длины пептиды и некоторое количество свободных аминокислот. Дальнейший гидролиз пептидов до свободных аминокислот осуществляется под влиянием группы ферментов - пептидаз. Продукты гидролиза белков всасываются в желудочно-кишечном тракте в основном в виде свободных аминокислот. Кинетика всасывания аминокислот в кишечнике в опытах in vivo и in vitro свидетельствует о том, что аминокислоты, подобно глюкозе, всасываются свободно с ионами натрия. 13.Пути использования аминокислот в организме: декарбоксилирование, образование биогенных аминов. В организме человека содержится около 100 г свободных аминокислот, которые образуют его аминокислотный фонд. Этот фонд постоянно пополняется за счёт поступления новых молекул аминокислот взамен тех, которые были использованы в метаболических процессах. Процесс отщепления карбоксильной группы аминокислот в виде СO2 получил название декарбоксилирования. Несмотря на ограниченный круг субстратов (аминокислот и их производных), подвергающихся декарбоксилированию в животных тканях, образующиеся продукты реакции (названные биогенными аминами) обладают сильным фармакологическим действием на множество физиологических функций человека и животных. Общая схема процесса декарбоксилирования аминокислот может быть представлена в следующем виде: R-CH(NH2)-COOH --> R-CH2-NH2 + CO2 В живых организмах открыто четыре типа декарбоксилирования аминокислот. 1.α -Декарбоксилирование.Продуктами реакции являются СО2 и биогенные амины: R-CH(NH2)-COOH --> R-CH2-NH2 + CO2 2.ω -Декарбоксилированне, характерное для микроорганизмов. Например, из аспарагиновой кислоты этим путем образуется α -аланин: НООС-СН2-CH(NH2)-СООН --> СН3-CH(NH2)-СООН + СО2 3.Декарбоксилирование, связанное с реакцией трансаминирования: Превращения аминокислот, вызванные деятельностью микроорганизмов кишечника, получили общее название гниения белков в кишечнике. Так, в процессе постепенного и глубокого распада серосодержащих аминокислот (цистина, цистеина и метионина) в кишечнике образуются сероводород (H2S) и метилмеркаптан (CH3SH). Диаминокислоты, в частности орнитин и лизин, подвергаются процессу декарбоксилирования с образованием протеиногенных аминов. Из ароматических аминокислот фенилаланина, тирозина и триптофана при аналогичном бактериальном декарбоксилировании образуются соответствующие биогенные амины: фенилэтиламин, парагидроксифенилэтиламин (или триптамин) и индолилэтиламин (триптамин). После всасывания эти продукты через воротную вену попадают в печень, где они подвергаются обезвреживанию В печени аминокислоты используются не только для синтеза собственных белков и белков плазмы крови, но также для синтеза ряда специфических азотсодержащих соединений Накопление биогенных аминов может отрицательно сказаться на физиологическом статусе и вызывать ряд серьезных нарушений в организме. Однако органы и ткани как и целостный организм располагают специальными механизмами обезвреживания биогенных аминов. 14.Дезаминирование, пути использования безазотистых остатков. Доказано существование четырех типов дезаминирования аминокислот (отщепление аминогруппы). Выделены соответствующие ферментные системы, катализирующие эти реакции, и идентифицированы продукты распада. Во всех случаях NH2-группа аминокислоты освобождается в виде аммиака: Помимо аммиака, продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты. Большая часть безазотистых остатков аминокислот превращается в пируват либо непосредственно (Ала, Сер), либо в результате более сложного пути, превращаясь вначале в один из метаболитов ЦТК. Затем в реакциях цитратного цикла происходит образование оксалоацетата, который превращается в фосфоенолпируват. Из фосфоенолпирувата под действием пируваткиназы образуется пируват. Пируват подвергается окислительному декарбоксилированию и превращается в ацетил-КоА, который окисляется в ЦТК до СО2 и Н2О с выделением энергии. Такой путь проходят преимущественно аминокислоты пищи. 15.Трансаминирование аминокислот. Диагностическое значение определения активности трансаминаз в организме. Под трансаминированием подразумевают реакции межмолекулярного переноса аминогруппы (NH2) от аминокислоты на α -кетокислоту без промежуточного образования аммиака. При добавлении к гомогенату мышц глутаминовой и пировиноградной кислот образуются α -кетоглутаровая кислота и аланин без промежуточного образования аммиака; добавление аланина и α -кетоглутаровой кислоты соответственно приводило к образованию пировиноградной и глутаминовой кислот: Аспартат + α -Кетоглутарат < --> ЩУК + Глутамат В сыворотке крови здоровых людей активность этих трансаминаз в среднем составляет 15 и 20 единиц по сравнению с десятками и сотнями тысяч единиц в других органах и тканях. Поэтому органические поражения при острых и хронических заболеваниях, сопровождающихся деструкцией клеток, приводят к выходу трансаминаз из очага поражения в кровь. При инфаркте миокарда уровень аспартатаминотрансферазы сыворотки крови уже через 3-5 ч после наступления инфаркта резко повышается, достигая 300-500 единиц. 16.Образование аммиака в организме и пути его обезвреживания. В организме человека подвергается распаду около 70 г аминокислот в сутки; при этом в результате дезаминирования, трансаминирования и окисления биогенных аминов освобождается большое количество аммиака, являющегося высокотоксичным соединением. Поэтому концентрация аммиака в организме должна сохраняться на низком уровне. Уровень аммиака в норме в крови не превышает 1-2 мг/л. Одним из путей связывания и обезвреживания аммиака в организме, в частности в мозге, сетчатке, почках, печени и мышцах, является биосинтез глутамина и аспарагина. 17.Биосинтез мочевины как основнои механизм предотвращения накопления аммиака. Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80-85% всего азота мочи. Кребс полагал, что каталитическую роль орнитина можно было бы объяснить, если бы существовал некий механизм для регенерации аргинина из орнитина, согласно уравнению: Орнитин + СO2 + 2NH3 --> Аргинин + 2H2O Ниже приведена, кроме того, суммарная реакция синтеза мочевины без учета промежуточных продуктов: СО2 + NH3 + 3АТФ + 2Н20 + Аспартат --> Мочевина + 2 АДФ + + АМФ + Фумарат + 2 Фн + ФФн
18.Синтез креатинина, креатина, креатин-фосфата и их значение ждя организма. Реакция синтеза креатина протекает в две стадии. Первая стадия – биосинтез гуанидинацетата – осуществляется в почках при участии глицин-амидинотранс-феразы: Вторая стадия синтеза креатина протекает в печени при участии гуанидинацетатметилтрансферазы: Креатин подвергается фосфорилированию с образованием креатин-фосфата, который после дефосфорилирования(необратимая реакция) превращается в креатинин, выделяющийся с мочой. Переваривание и всасывание нуклеопротеидов. Образование мочевой кислоты и патологии, связанные с ее синтезом Переваривание нуклеопротеинов и всасывание продуктов их распада осуществляются в пищеварительном тракте. Под влиянием ферментов желудка, частично соляной кислоты, нуклеопротеины пищи распадаются на полипептидыи нуклеиновые кислоты; первые в кишечнике подвергаются гидролитическому расщеплению до свободныхаминокислот. Распад нуклеиновых кислот происходит в тонкой кишке в основном гидролитическим путем под действием ДНК- и РНКазы панкреатического сока. Продуктами реакции при действии РНКазы являются пуриновые и пи-римидиновые мононуклеотиды. В отношении дальнейшей судьбы мононуклеотидов существует два предположения. Считают, что мононуклеотиды в кишечнике под действием неспецифических фосфатаз (кислой и щелочной), которые гидролизируют фосфоэфирную связь мононуклеотида («нуклеотидазное» действие), расщепляются с образованием нуклеозидов и фосфорной кислоты и в таком виде всасываются. Согласно второму предположению, мононуклеотиды всасываются, а распад их происходит в клетках слизистой оболочки кишечника. Образовавшиеся при гидролизе пуриновые нуклеозиды – аденозин и гуано-зин – подвергаются ферментативному распаду в организме животных вплоть до образования конечного продукта – мочевой кислоты, которая выводится с мочой из организма.
Повышенное выделение мочевой кислоты наблюдается при лейкемии, полицитемии, гепатитах и подагре. Содержание мочевой кислоты в моче повышается также при приеме ацетилсалициловой кислоты и ряда стероидных гормонов. Наряду с мочевой кислотой в моче всегда содержится небольшое количество пуринов экзо, и эндогенного происхождения.
20.Механизм возникновения наследственных нарушений обмена аминокислот (фенилкетонурия, альбинизм, алкаптонурия, болезнь паркинсона) Возникновение и дальнейшее развитие специфического патологического синдрома при таких заболеваниях обусловлено полным или частичным отсутствием активности определенных ферментов: организм либо теряет способность синтезировать данный фермент, либо образуется недостаточное количество его, либо синтезируется аномальный фермент, отличающийся по структуре от нативного. Следствием такого врожденногодефекта обмена является накопление в тканях нормальных промежуточных или побочных (неспецифических) продуктов обмена, оказывающих токсическое влияние на организм и в первую очередь на ЦНС. Этим, пожалуй, объясняется тот факт, что в основном заболевают дети в раннем возрасте, у которых затем развиваются специфические расстройства психической деятельности. Фенилкетонурия (фенилпировиноградная олигофрения) развивается как результат потери способности организмасинтезировать фенилаланин-4-монооксигеназу, катализирующую превращение фенилаланина в тирозин. Характерные особенности болезни – резкое замедление умственного развития ребенка, а также экскреция с мочойбольших количеств фенил-пировиноградной кислоты (до 1–2 г/сут) и фенилацетилглутамина (до 2–3 г/сут). Решающим доказательством метаболического блока при фенил-кетонурии являются данные о накоплениифенилаланина в тканях. Так, количество его в крови может достигать 600 мг/л (в норме 15 мг/л), вцереброспинальной жидкости – 80 мг/л (в норме 1, 5 мг/л). Развитие болезни можно предотвратить, если значительно снизить прием фенилала-нина с пищей с самого рождения ребенка. Алкаптонурия характеризуется экскрецией с мочой больших количеств (до 0, 5 г/сут) гомогентизиновой кислоты, окисление которой кислородом воздуха придает моче темную окраску. В далеко зашедших случаях развиваются охроноз, наблюдаются отложение пигмента в тканях и потемнение носа, ушей и склеры. Эта болезнь известна с девнейших времен, однако только в 1962 г. были получены доказательства, что метаболический дефект при алкаптонурии связан с врожденным отсутствием в печени и почках оксидазы гомогентизиновой кислоты. Альбинизм – врожденное отсутствие пигментов в коже, волосах и сетчатке. Метаболический дефект связан с потереймеланоцитами способности синтезировать тирозиназу – фермент, катализирующий окисление тирозина в диоксифенилаланин и диоксифенилаланинхинон, являющихся предшественниками меланина. Предположение о блокировании процесса полимеризации меланина при альбинизме не подтвердилось.
Гормоны щитовидной железы. Щитовидная железа состоит из множества особых полостей – фолликулов, заполненных вязким секретом – коллоидом. В состав коллоида входит особый йодсодержащий гликопротеин с высокой мол. массой – порядка 650000 (5000 аминокислотных остатков). Этот глико-протеин получил название йодтиреоглобулина. Он представляет собой запасную форму тироксина и трийодтиронина – основных гормоновфолликулярной части щитовидной железы. Помимо этих гормонов (биосинтез и функции которых будут рассмотрены ниже), в особых клетках – так называемых парафолликулярных клетках, или С-клетках щитовидной железы, синтезируется гормон пептидной природы, обеспечивающий постоянную концентрацию кальция в крови. Он получил название «кальцитонин». Биологическое действие кальцитонина прямо противоположно эффектупаратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Таким образом, постоянство уровня кальция в крови человека и животных обеспечивается главным образом паратгормоном, кальцитриолом и кальцитонином, Химическая природа гормонов фолликулярной части щитовидной железы выяснена в деталях сравнительно давно. Считается установленным, что все йодсодержащие гормоны, отличающиеся друг от друга содержанием йода, являются производными L-тиронина, который синтезируется в организме из аминокислоты L-тирозина.
Биологическое действие гормонов щитовидной железы распространяется на множество физиологических функцийорганизма. В частности, гормоны регулируют скорость основного обмена, рост и дифференцировку тканей, обменбелков, углеводов и липидов, водно-электролитный обмен, деятельность ЦНС, пищеварительного тракта, гемопоэз, функцию сердечнососудистой системы, потребность в витаминах, сопротивляемость организма инфекциям и др Гипофункция щитовидной железы в раннем детском возрасте приводит к развитию болезни, известной в литературе как кретинизм. Помимо остановки роста, специфических изменений кожи, волос, мышц, резкого снижения скорости процессов обмена, при кретинизме отмечаются глубокие нарушения психики; специфическое гормональное лечение в этом случае не дает положительных результатов. Недостаточная функция щитовидной железы в зрелом возрасте сопровождается развитием гипотиреоидного отека, или микседемы (от греч. myxa – слизь, oedemo – отек). Это заболевание чаще встречается у женщин и характеризуется нарушением водно-солевого, основного и жирового обмена. У больных отмечаются слизистый отек, патологическое ожирение, резкое снижение основного обмена, выпадение волос и зубов, общие мозговые нарушения и психические расстройства. Кожа становится сухой, температура тела снижается; в крови повышено содержание глюкозы. Гипотиреоидизм сравнительно легко поддается лечению препаратами щитовидной железы. Следует отметить еще одно поражение щитовидной железы – эндемический зоб. Болезнь обычно развивается у лиц, проживающих в горных местностях, где содержание йода в воде и растениях недостаточно. Недостаток йодаприводит к компенсаторному увеличению массы ткани щитовидной железы за счет преимущественного разрастаниясоединительной ткани, Лечение сводится к обогащению продуктов питания, в частностиповаренной соли, неорганическим йодом. Повышенная функция щитовидной железы (гиперфункция) вызывает развитие гипертиреоза, известного в литературе под названием «зоб диффузный токсический» (болезнь Грейвса, или базедова болезнь). Резкое повышение обмена веществ сопровождается усиленным распадом тканевых белков, что приводит к развитию отрицательного азотистого баланса. Наиболее характерным проявлением болезни считается триада симптомов: резкое увеличение числа сердечных сокращений (тахикардия), пучеглазие (экзофтальм) и зоб, т.е. увеличенная в размерах щитовиднаяжелеза; у больных отмечаются общее истощение организма, а также психические расстройства. При гиперфункции щитовидной железы и, в частности, токсическом зобе показано оперативное удаление всей железы или введение
Белки крови и их роль. Из 9–10% сухого остатка плазмы крови на долю белков приходится 6, 5–8, 5%. Используя метод высаливания нейтральными солями, белки плазмы крови можно разделить на три группы: альбумины, глобулины и фибриноген. Нормальное содержание альбуминов в плазме крови составляет 40–50 г/л, глобулинов – 20–30 г/л, фибриногена – 2, 4 г/л. Плазма крови, лишенная фибриногена, называется сывороткой. Синтез белков плазмы крови осуществляется преимущественно в клетках печени и ретикулоэндотелиальной системы. Физиологическая роль белков плазмы крови многогранна. 1. Белки поддерживают коллоидно-осмотическое (онкотическое) давление и тем самым постоянный объем крови. 2. Белки плазмы принимают активное участие в свертывании крови.. 3. Белки плазмы в известной мере определяют вязкость крови, 4. Белки плазмы принимают участие в поддержании постоянного рН крови, так как составляют одну из важнейшихбуферных систем крови. 5. Важна также транспортная функция белков плазмы крови: соединяясь с рядом веществ (холестерин, билирубин и др.),. 6. Белки плазмы играют важную роль в процессах иммунитета (особенно это касается иммуноглобулинов). 7. В результате образования с белками плазмы недиализируемых комплексов поддерживается уровень катионов вкрови. 8. Наконец, белки плазмы крови могут служить резервом аминокислот. Альбумины. На долю альбуминов приходится более половины (55–60%)белков плазмы крови человека. Мол. масса альбумина около 70000. Сывороточные альбумины сравнительно быстро обновляются (период полураспада альбуминов человека 7 дней). Благодаря высокой гидрофильности, особенно в связи с относительно небольшим размером молекул и значительнойконцентрацией в сыворотке, альбумины играют важную роль в поддержании онкотического давления крови. Известно, что концентрация альбуминов в сыворотке ниже 30 г/л вызывает значительные изменения онкотического давлениякрови, что приводит к возникновению отеков. Альбумины выполняют важную функцию транспорта многих биологически активных веществ (в частности, гормонов). Они способны связываться с холестерином, желчными пигментами. Значительная часть кальция в сыворотке крови также связана с альбуминами. Глобулины. Сывороточные глобулины при высаливании нейтральными солями можно разделить на 2 фракции – эуглобулины и псевдоглобулины. Фракция эуглобулинов в основном состоит из γ -глобулинов, а фракция псевдоглобулинов включает α -, β - и γ -глобулины, которые при электрофорезе, особенно в крахмальном или полиакриламидном геле, способны разделяться на ряд подфракций. α - и β -Глобулиновые фракции содержатлипопротеины, а также белки, связанные с металлами. Большая часть антител, содержащихся в сыворотке, находится во фракции γ -глобулинов. При снижении уровня белков этой фракции резко понижаются защитные силыорганизма.
Гиперпротеинемия – увеличение общего содержания белков плазмы. Диарея у детей, рвота при непроходимости верхнего отдела тонкой кишки, обширные ожоги могут способствовать повышению концентрации белков в плазме крови. Иными словами, потеря воды организмом, а следовательно, и плазмой приводит к повышению концентрациибелка в крови (относительная гиперпротеинемия). Гипопротеинемия, или уменьшение общего количества белка в плазме крови, наблюдается главным образом при снижении уровня альбуминов. Выраженная гипопротеинемия – постоянный и патогенетически важный симптом нефротического синдрома. Содержание общего белка снижается до 30–40 г/л. Гипопротеинемия наблюдается также при поражении печеночных клеток (острая атрофия печени, токсический гепатит и др.). Кроме того, гипопротеинемия может возникнуть при резко увеличенной проницаемости стенок капилляров, при белковой недостаточности (поражение пищеварительного тракта, карцинома и др.). Следовательно, можно считать, что гиперпротеинемия, как правило, связана с гиперглобулинемией, а гипопро-теинемия – с гипоальбуминемией.
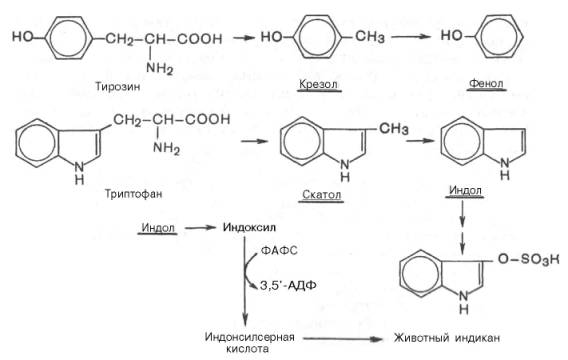
Образование токсинов в толстом кишечнике из аминокислот. превращения аминокислот, вызванные деятельностью микроорганизмов кишечника, получили общее название «гниение белков в кишечнике». Из ароматических аминокислот: фенилаланин, тирозин и триптофан – при аналогичном бактериальномдекарбоксилировании образуются соответствующие амины: фенилэтиламин, параоксифенилэтиламин (или тира-мин) и индолилэтиламин (триптамин). Кроме того, микробные ферменты кишечника вызывают постепенное разрушение боковых цепей циклических аминокислот, в частности тирозина и триптофана, с образованием ядовитых продуктов обмена – соответственно крезола и фенола, скатола и индола.
36Выяснение влияния желчи на активность липазы. Липаза-малоспецифический фермент, который действует на многие жиры при рН 9, 0. Липаза гидролитически расщепляет жиры и в первую очередь эфирную связь в а-положении. ПРИНЦИП МЕТОДА: скорость действия липазы в отдельных порциях жира молока модно узнать по количеству жирных кислот, образующихся при гидролизе жира за определенный промежуток времени. Кол-во жирных к-т определяют титрованием щелочью в присутствии фенолфталеина.результаты выражают в мл титрованного раствора щелочи и строят график. ХОДЖ в тва стакана наливают молоко и панкреатин. В 1 ст приливают воду, а в другой желчь и перемешивают. Потом добавляют фенолфталеин в каждую и титруют 0, 05% рс-м едкого натра до слабо-розовой окраски. Помещают в термостат при 38. и через каждые 10мин отбирают по 1 мл и титруют рс-м едкого натра в присутствии фенолфталеина. Повторяют 5 6 раз. И строят график. 37действие фосфолипаз поджелудочной железы. В поджелудочной железе и ее соке содержится несколько фосфолипаз-ферментов, ускоряющих гидролиз фосфолипидов, в частности лецитина. ПРИНЦИП МЕТОДА: об активности фосфолипазподж.железы судят по появлению свободной фосфорной кислоты, способной образовывать желтый осадок при нагревании с молибдатом аммония. ХОД: в 2 пробирки нали-ют суспензии яичного желтка. В 1-ю доб-ют панкреатин, а во 2-ю воду. Обе пр-ки помещают в термостат при 38 на 30 мин. После, в обе пр-ки наливают молибденового реактива и нагревают их на пламени горелки и охлаждают водой. 38эмульгирование жиров. Эмульгирование жиров в кишечнике осуществляется при участии солей желчных кислот. Основное переваривание жиров происходит в тонком кишечнике. Соли желчных кислот адсорбируются в присутствии небольших количеств свободных жирных кислот и моноглицеридов на поверхности капелек жира в виде тончайшей пленки, препятствующей слиянию этих капелек. Кроме того, соли желчных кислот, уменьшая поверхностное натяжение на границе раздела фаз вода — жир, способствуют дроблению больших капелек жира на меньшие. Создаются условия для образования тонкой и устойчивой жировой эмульсии с частицами диаметром 0, 5 мкм и меньше. В результате эмульгирования резко увеличивается поверхность капелек жира, что увеличивает площадь их взаимодействия с липазой, т.е. ускоряет ферментативный гидролиз, а также всасывание. Всасывание жиров, как и других липидов, происходит в проксимальной части тонкой кишки. Тонко эмульгированные жиры могут частично проникать через стенку кишечника без предварительного гидролиза.. Однако основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные кислоты и моноглицериды. Всасывание этих соединений происходит при участии желчи. Жирные кислоты и моноглицериды образуют с компонентами желчи жировые мицеллы, которые могут связывать дополнительные количества неэтерифицированногохолестерина, а также жирорастворимые витамины (A, D, Е и К), Жировые мицеллы рассматривают как своеобразный комплекс липидов и продуктов их распада с желчными кислотами и другими компонентами желчи в кишечнике.обеспечивающий растворимость липидов и продуктов их распада в водной среде (так называемаямицеллярнаясолюбилизация липидов) и их всасывание кишечной стенкой. Исключительно важное значение при этом имеет гепатобилиарная циркуляция желчных кислот, обеспечивающая всасывание больших количеств жирных кислот и моноглицеридов (50—70, а иногда и более 100 г) при относительно невысоком общем количестве желчных кислот в организме (2, 8—3, 5 г). 39 определение свободных жирных кислот. в крови сод-ся свободных жирных кислот 640-880мкмоль/л. ПРИНЦИП МЕТОДА: медные соли жирных кислот способны образовывать с диэтилдитиокарбаматом натрия окрашенные комплексные соединения, интенсивность окраски которых пропорциональна концентрации свободных жирных кислот. ХОД: в 1-ю пр-ку вносят сыворотку крови, а в другую пальмитиновую кислоту в хлороформе. В обе пробирки доб-ют хлороформ и медного реактива. Пробирки закрывают и встряхивают в течении 3 мин. Содержимое центрифугируют при 3000об/мин в течение 15 мин. Смесь в пробирках разд-ся на три слоя: хлороформ, белок, вода. водную фазу удаляют, белковую пленку сдвигают на стенки пробирок =, а хлороформный слой переносят в пробирки и к это слою доб-ют р-р диэтилдитиокарбамата натрия в бутаноле и перемешивают. Содержимое своб.жир.кислотрассч-ют по формуле Е пробы х 1000/Е стандарта х 0, 5 мкмоль/л |
Последнее изменение этой страницы: 2017-04-13; Просмотров: 684; Нарушение авторского права страницы




 4. Декарбоксилирование, связанное с реакцией конденсации двух молекул:
4. Декарбоксилирование, связанное с реакцией конденсации двух молекул: 

 Для клинических целей наибольшее значение имеют две трансаминазы - аспартатаминотрансфераза и аланинаминотрансфераза, катализирующие соответственно следующие обратимые реакции:
Для клинических целей наибольшее значение имеют две трансаминазы - аспартатаминотрансфераза и аланинаминотрансфераза, катализирующие соответственно следующие обратимые реакции:  Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80-85% всего азота мочи.
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80-85% всего азота мочи.