|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ПАРАСИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
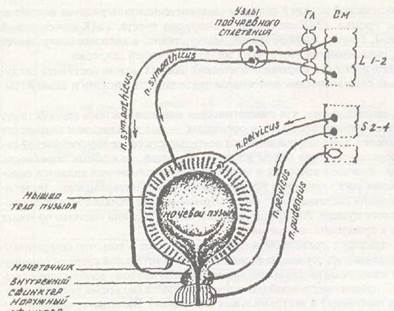
Центральные (преганглионарные) нейроны парасимпатической нервной системы расположены в среднем, продолговатом мозге и в люмбосакральном отделе спинного мозга. В среднем мозге расположены два парасимпатических ядра, относящихся к Ш паре — ядро Якубо-вича-Вестфаля-Эдингера (иннервация сфинктера зрачка) и часть его — ядро Перлеа, иннер-вирующего ресничную мышцу глаза. В продолговатом мозгу имеются парасимпатические ядра VII, IX, X пар черепно-мозговых нервов. Парасимпатическое ядро VII пары шшервиру-ет слизистые железы полости носа, слезную железу, а через chorda tympani — подъязычную и подчелюстную слюнные железы. Парасимпатическое ядро IX пары иннервирует околоушную железу. Парасимпатическое ядро X пары (вагуса) — одно из самых мощных. Оно иннервирует органы шеи, грудной и брюшной полостей (сердце, легкие, желудочно-кишечный тракт). В пояснично-сакральном отделе спинного мозга расположены парасимпатические нейроны, которые образуют центры мочеиспускания, дефекации, эрекции. Распространенность влияния парасимпатического отдела более ограничена, чем симпатического. Почти все сосуды тела не имеют парасимпатических волокон. Исключение — сосуды языка, слюнных желез и половых органов. Как и симпатическая система, парасимпатическая имеет преганглионарные нейроны, аксоны которых идут к органу (постганглионарные волокна). Ганглии парасимпатической нервной системы находятся, как правило, в толще органа (интрамуральные ганглии), поэтому преганглионарные волокна — длинные, а постганглионарные — короткие. С органом контактирует постганглионарное волокно. Оно либо непосредственно взаимодействует с клетками этого органа (ГМК, железы), либо опосредованно через метасимпатическую нервную систему. В преганглионарных волокнах парасимпатической нервной системы медиатором является ацетилхолин, он взаимодействует на постсинаптической мембране постганглионарно-го нейрона с Н-холинорецепторами, которые блокируются ганглиоблокаторами. Следова-
тельно, передача возбуждения с пре-ганглионарного волокна на постганг-лионарный нейрон в парасимпатической системе происходит так же, как и в симпатической нервной системе. В окончаниях пост-гаиглионарных волокон парасимпатической нервной системы, в отличие от симпатической, выделяется ацетил-холин, а рецепторы, расположенные на постсинаптичес-кой мембране органа (или нейрона ме-тасимаатической системы) — это М-холинорецепторы (мус каринчу встви-тельные холиноре-цепторы), которые блокируются веществами типа атропина. Какие эффекты оказывает парасимпатическая система? На сердце — угнетение частоты, силы, проводимости и возбудимости, ГМК бронхов — активация (это приводит к сужению бронхов), секреторные клетки трахеи и бронхов — активация, ГМК и секреторные клетки ЖКТ — активация, сфинктеры ЖКТ, сфинктеры мочевого пузыря — расслабление, дструс-сор (мышца мочевого пузыря) — активация, сфинктер зрачка — активация, ресничная мышца глаза — активация (повышается кривизна хрусталика, усиливается преломляющая способность глаза), повышение кровенаполнения сосудов половых органов, активация слюноотделения, повышение секреции слезной жидкости. В целом, возбуждение парасимпатических волокон приводит к восстановлению гомеостаза, т. е. к трофотропному эффекту. ВЫСШИЕ ВЕГЕТАТИВНЫЕ ЦЕНТРЫ Гипоталамус является одним из важнейших образований мозга, участвующих в регуляции активности нейронов парасимпатической и симпатической нервной системы. Гипоталамус представляет собой скопление более чем 32 пар ядер. Существует большое разнообразие в классификации ядер гипоталамуса. 1. Деление на эрготропные и трофотропные ядра (классификация Гесса) — ядра, кото 2. Деление на симпатические и парасимпатические ядра — полагают, что в передних симпатической нервной системы, в задних же отделах — ядра, вызывающие активацию симпатического отдела ВНС. Более академично, очевидно, деление гипоталамических ядер по их топографическим признакам. В настоящее время большинство авторов разделяет весь гипоталамус на 4 области или группы ядер: 1) преоптическую, 2) переднюю, 3) среднюю (туберальная, или группа ядер срединного бугра) и 4) заднюю группу ядер. В каждой из этих областей выделяют отдельные ядра. Перечислим основные ядра, знание которых облегчит восприятие многих деталей, связанных с функцией гипоталамуса. Таблица 2.
В целом в этих ядрах локализуются центры, участвующие в вегетативной регуляции, а также нейросекреторные нейроны, осуществляющие секрецию нейрогормонов и веществ типа либеринов и статинов. Рассмотрим подробнее функции гипоталамуса и свойства отдельных ядер. Гипоталамус имеет обширные афферентные и эфферентные связи. Все рецепторы в конечном итоге доставляют свою информацию и в ядра гипоталамуса. Функции гипоталамуса. Полагают, что в гипоталамусе имеются ядра, которые преимущественно активируют либо парасимпатические нейроны ствола и спинного мозга, либо симпатические нейроны спинного мозга. Соответственно их принято называть трофотроп-ными и эрготропными ядрами. Они расположены соответственно в передних и задних отделах гипоталамуса. Однако следует помнить, что нет такой четкой локализации, какую бы хотелось иметь с позиций усвоения материала. Ядра гипоталамуса являются высшими вегетативными центрами. Существуют более конкретные представления о роли гипоталамуса в регуляции вегетативных функций. В частности, известно, что в гипоталамусе имеются центры терморегуляции. Среди ядер передней группы имеются нейронные скопления, которые регулируют процесс отдачи тепла, поэтому эти скопления называются центром физической терморегуляции; среди ядер задней группы гипоталамуса имеются скопления нейронов, ответственных за процесс теплопродукции — это центр химической терморегуляции. Ког- да указанные образования разрушаются, например, в эксперименте, то животное лишается возможности переносить перенагревание или переохлаждение соответственно. В гипоталамусе находятся центры регуляции водного и солевого обмена. В частности, в передней группе ядер гипоталамуса среди нейронов паравентрикулярного и супраоптическо-го ядер имеются нейроны, участвующие в этом процессе, в том числе за счет продукции антидиуретического гормона, а среди ядер средней группы гипоталамуса находится центр жажды, обеспечивающий поведение животного или человека, направленное на прием воды (нормализацию водно-солевого обмена). В гипоталамусе находятся центры белкового, углеводного и жирового обменов, центры регуляции сердечнр-сосудистой системы, проницаемости сосудов и тканевых мембран, морфообразования, эндокринных функций (желез), деятельности желудочно-кишечного тракта (в том числе голода, который локализован в латеральном гипотала-мнческом ядре, где и центр жажды, а в вентромедиальном ядре — расположен центр насыщения), центры регуляции мочеотделения, регуляции сна и бодрствования, полового, эмоционального поведения, центры, участвующие в процессах адаптации организма. Относительно некоторых ядер гипоталамуса остановимся подробнее. 1. Супрахиазматическое ядро — ядро передней группы гипоталамуса. Полагают, что его Нейроны супрахиазматического ядра обладают свойством автоматик и поэтому являются внутренними «часами» организма. За счет наличия прямых связей этого ядра с сетчаткой глаза (ретиногипоталамические волокна) ритм нейронов этого ядра приурочен к изменению освещенности (день-ночь), в связи с чем многие циркадные ритмы зависят от времени суток. Нейроны этого ядра получают воздействия от центров, модулирующих биоритм. Например, сюда поступают сигналы от серотонинергических, адренергических, дофами-нергических и холинергических нейронов ствола мозга. Особенно важна информация, идущая от серотонинергических нейронов шва: серотонин может тормозить активность нейронов супрахиазматического ядра. Таким образом, изменения, возникающие в отдельных структурах мозга, могут за счет влияния на водитель циркадного ритма существенно менять сам характер этого ритма. 2. Супраоптнческое и паравентрнкулярное ядра. Нейроны этих образований помимо уча Нейроны супраоптического и паравентрикулярного ядер помимо секреции этих гормонов способны управлять выделением этих же гормонов — управляют процессом синапти-ческой передачи, где в качестве медиатора фактически выступает окситоцин или АДГ. Регуляция этого процесса идет за счет приходящего от тела нейрона потенциала действия, который вызывает деполяризацию пресинаптической мембраны и это (как в обычном синапсе) приводит к вхождению в синапс ионов кальция, что вызывает экзоцитоз и выделение гормона-медиатора. Часть нейронов гипоталамуса продуцирует пептидные гормоны, которые получили название либеринов (рилизинг-гормонов или освобождающих гормонов) и статинов, гормонов, которые тормозят выделение других гормонов. Область, в которой локализованы эти нейроны, называется гипофизотропной областью, а нейроны, продуцирующие либерины или статины — тубероинфундибулярные нейроны или по «имени» соответствующего гормона. Например, кортиколиберинпродуцирующий нейрон. Оказалось, что нейроны, продуцирующие либерины и статины, локализованы во многих отделах гипоталамуса, сосредо- точены в различных ядрах, в том числе в супраоптическом и паравентрикулярном ядрах. Например, кортиколиберин синтезируется в паравентрикулярном ядре (это основное место его продукции), в супраоптическом, супрахиазматическом, аркуатном, дорсомедиаль-ном и вентромедиальном ядрах. Все эти ядра содержат нейроны, аксоны которых идут в срединное возвышение: здесь расположены капилляры верхней гипофизарной артерии, куда и вливаются либерины и статины. С током крови либерины и статины достигают передней доли гипофиза и вызывают изменение продукции соответствующего гормона аденогипо-физа. В настоящее время известно, что в гипоталамусе вырабатываются 6 либеринов и 3 статина. Либерины: тиролиберин, люлиберин, кортиколиберин, соматолиберин, рилизинг-гормон пролактина и рнлизинг-гормон меланоцитстимулирующего гормона. Статины: со-матостатин, ингибитор выделения МСГ и ингибитор выделения ПРЛ. Таким образом, благодаря обширным связям гипоталамуса с различными структурами мозга, за счет продукции гормонов и нейросекреции гипоталамус участвует в регуляции многих функций организма через гуморальное звено регуляции, изменяя продукцию гормонов гипофиза. Гшготаламо-гипофизарные связи, которые, как видно из сказанного, имеют два варианта — связь через аксоны с нейрогипофизом и через систему портальных сосудов с передним гипофизом — играют очень важную роль в жизнедеятельности организма, в связи с чем им уделяется такое большое внимание. Гипоталамус является компонентом лимбической системы. В нее входят многие структуры старой и древней коры больших полушарий, в том числе гиппокамп, миндалевидный комплекс. Гипоталамус является центральной структурой лимбической системы: именно благодаря гипоталамусу все эмоциональные реакции, которые реализуются с участием лимбической системы мозга, приобретают конкретную вегетативную и эндокринную окраску. МОНОАМИНЕРГИЧЕСКАЯ СИСТЕМА Моноаминергическую систему в последние годы относят к центрам, осуществляющим регуляцию вегетативных функций. Эта система объединяет в себе скопление нейронов ствола мозга, аксоны которых идут в составе медиального пучка переднего мозга и достигают практически всех структур мозга — в том числе гипоталамуса, таламуса, базальных ганглиев, коры (лимбической коры и новой коры). В систему входят три типа нейронов: норадренер-гические, дофаминергические и серотонинергические. Тела норадренергических нейронов расположены в продолговатом мозге и в мосту, особенно их много в голубом пятне. Эти нейроны оказывают тормозящее влияние на структуры ЦНС. Тела дофаминергических нейронов лежат в среднем мозге, в основном, в черной субстанции. Аксоны их, помимо связи с базальными ганглиями, влияют и на другие структуры мозга. Серотонинергические нейроны локализованы в продолговатом мозгу — в основном, в ядрах срединного шва, в мосту и в нижних отделах среднего мозга. Считается, что моноаминергическая система (НА, серотонин и дофамин — это моноамины) играет важную роль в создании эмоций и вегетативного обеспечения эмоционального состояния. Это осуществляется за счет воздействия нейронов моноаминергической системы одновременно на структуры лимбической системы и на ядра гипоталамуса, которые управляют вегетативными нейронами ствола мозга и спинного мозга. Норадреналин создает условия для появления у человека положительных эмоций — благодаря норадрена-лину в основном функционируют центры удовольствия. Поэтому у больных, принимающих вещества, которые блокируют накопление в нейронах норадреналина (например, резерпин), может развиваться депрессивное состояние как результат дефицита норадреналина в нейронах. Дофамин, подобно норадреналину г способствует созданию положительных эмоций. При чрезмерной выработке дофамина, как полагают, создаются условия для шизофрении. Не случайно среди лекарственных средств, применяемых для лечения шизофрении, имеются вещества, подавляющие синтез дофамина. Серотонин играет роль в создании отрицатель- ных эмоций — при этом активируются центры избегания. Транквилизаторы за счет подавления выработки серотонина, как известно, способны снять отрицательные эмоции. Итак, моноаминергическая система играет определенную роль в создании эмоций и в изменении деятельности внутренних органов за счет воздействия на многие высшие вегетативные центры, в том числе — гипоталамус. МОЗЖЕЧОК По мнению Леона Абгаровича Орбели, мозжечок имеет важное значение в регуляции вегетативных функций организма. По Орбели, мозжечок играет, как и симпатическая нервная система, адаптационно-трофическую роль, т. е. способствует активации всех резервов организма для выполнения мышечной работы. Будучи одним из важнейших центров, участвующих в регуляции двигательной (мышечной) активности, мозжечок должен принимать участие и в регуляции вегетативного обеспечения мышечной активности. Итак, мозжечок не только координирует произвольную мышечную активность с процессами поддержания равновесия, но и одновременно способствует согласованию (координации) вегетативных функций в этот период. При удалении мозжечка, однако, не наступает смерть (мозжечок не является жизненно необходимым образованием). Но при этом, как показывают эксперименты на животных, нарушается вегетативная регуляция. Скорее всего, мозжечок влияет на возбудимость вегетативных нервных центров и тем самым способствует адаптации организма к выполнению двигательных актов. С этих позиций мозжечок можно рассматривать как посредник между вегетативной нервной системой и соматической нервной системой. Показано, что мозжечок влияет на деятельность сердца, меняет величину артериального давления, регионарный кровоток (при удалении мозжечка снижается интенсивность кровотока в красных мышечных волокнах без существенного изменения кровотока в белых мышечных волокнах), влияет на глубину и частоту дыхания, на моторную, секреторную, всасывательную функцию ЖКТ, на процессы желчеобразования, влияет на тонус мышц мочевого пузыря. При повреждении мозжечка нарушается углеводный, белковый и минеральный обмен, нарушается энергообразование и процессы терморегуляции, нарушаются процессы кроветворения. Мозжечок имеет отношение и к регуляции процессов репродукции. Так, например, у беременных кошек электростимуляция мозжечка вызывает активацию матки. При удалении мозжечка у беременной кошки пролонгируется беременность, а рожденное потомство — нежизнеспособно. У собак после удаления мозжечка нарушается половое созревание и такие собаки не способны иметь потомство. Показано, что для нормального течения беременности необходимо, чтобы у животного была сохранена хотя бы часть мозжечка. Итак, приведенные сведения показывают, что мозжечок играет важную роль в процессах регуляции вегетативных функций. Будущие исследования должны уточнить и конкретизировать механизмы, посредством которых мозжечок влияет на вегетативные центры ствола мозга и спинного мозга. |
Последнее изменение этой страницы: 2017-05-04; Просмотров: 1492; Нарушение авторского права страницы