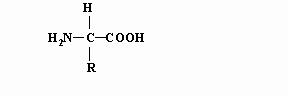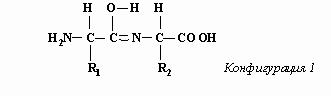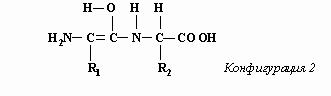|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Как проводили биологическую эволюцию: виды-инкубаторы и виды-выводки.
Материалистическая наука полагает, что всё на свете происходит без сверхъестественных вмешательств. В частности, совершенно естественно происходит и биологическая эволюция, причём новые биологические виды появляются в результате совершенно естественного отбора, а не чего-то там ещё.
По ходу того, как эта концепция деревенела в массовом сознании, оказывались несостоятельными аргументы и факты, на которых она была основана, а количество противоречащих ей аргументов и фактов росло, как снежный ком. Так, селекционеры скрестили всех, кого смогли, но за всю историю искусственной селекции не вывели ни одного нового вида. Палеонтологи докопались до того, что новые виды возникали внезапно, без переходных форм, которые требовались по логике естественной эволюции. Причём в истории было несколько периодов, когда видообразование имело взрывообразный характер. Биологи теряли дар речи перед фактами грандиозных качественных скачков в морфологии и физиологии организмов, а также перед появлением у них изумительных приспособлений, для которых в принципе не могло быть прототипов. Биохимики не обнаружили никаких восходящих эволюционных линий на уровне строения биомолекул, а генетики, к своему ужасу – на уровне составов геномов. Инженеры скромно намекнули: чтобы в системе накапливались изменения в некотором определённом направлении, мощность направляющего воздействия должна хоть немного превышать мощность случайных воздействий. В результате только случайных мутаций никакие эволюционные поползновения невозможны! Математики всё же, узнав про генетический код, оценили время, за которое, скажем, лапа могла превратиться в ласт в результате счастливой последовательности случайных мутаций, и получили сумасшедшие триллионы лет. Причём, почти всё это время кто-то ведь должен охранять и кормить промежуточных бедолаг, не имеющих возможности ни бегать, ни плавать. Ну, а физики, проводящие радиоактивную датировку образцов пород и окаменелостей, вплоть до тех, чей возраст по естественно-эволюционным раскладкам составляет сотни миллионов лет, исправно находят в них изотоп С14, который должен был в них практически полностью иссякнуть где-то за 20 тысяч лет. На этом парад несуразностей не заканчивается. Здравомыслящие люди давно понимают, что естественным отбором нельзя объяснить эволюцию, ибо естественный отбор может лишь устранять биологические виды с лица земли, но он не может их создавать. Действительно, благодаря естественному отбору “одни выживают, а другие вымирают”. Но заметим, что прежде и “те”, и “другие” жили на белом свете. Значит, если действует только естественный отбор, то число биологических видов неуклонно сокращается. И если в результате этого неуклонного сокращения современное видовое многообразие составляет, по самым скромным подсчётам, два с половиной миллиона, то пусть сторонники естественной эволюции ответят на смешные вопросы о том, сколько же видов было на старте эволюции, и, главное, откуда они там взялись.
Впрочем, эти сторонники ради такого случая состряпали вспомогательную теорию дивергенции, т.е. расщепления одного биологического вида на несколько. Причём происходит это, якобы, тоже совершенно естественным путём. Увы! Теоретики дивергенции, похоже, подзабыли главное свойство, по которому биологические виды обособлены друг от друга. Так мы напомним. Это свойство следующее: жизнеспособное и плодовитое потомство получается лишь от самцов и самок одного и того же вида. При этом, разумеется, потомство принадлежит к тому же виду, что и родители. Сразу возникает детский вопрос: “Дяденьки академики, а откуда берутся дети, принадлежащие к новым видам? Может, и вправду их приносят аисты? ” На это академики дают разъяснения. Мол, аисты ни при чём, а просто мутации копятся-копятся, и вот наступает знаменательный день, когда некая мамаша производит на свет детёныша вполне нового вида. А где же доказательства того, что он уже нового вида? А это ясно по определению: когда он вырастет, ему не с кем будет спариться, чтобы произвести жизнеспособное и плодовитое потомство. И на этом вся дивергенция бесславно закончится.
Как ни крути, а единственный шанс, который имеется у детёнышей нового вида – это их массовое и почти одновременное рождение. Мы полагаем, что именно по такому сценарию начинается история нового вида. Причём, массовые удачные зачатия и благополучное эмбриональное развитие новичков абсолютно невозможны в результате наложения случайностей. События такого рода могут быть лишь результатом тщательно подготовленной и великолепно проведённой операции. Такая операция с материалистической точки зрения является сверхъестественным вмешательством, поскольку она осуществляется Программистами, работающими на программном уровне реальности. Обрисуем в самых общих чертах программные манипуляции, которые необходимо проделать для созидания нового биологического вида.
Этот новый вид мы будем называть видом-выводком, а вид, из которого он выводится – видом-инкубатором. Представители вида-выводка могут весьма сильно отличаться от представителей вида-инкубатора, особенно при созидании нового таксона крупного ранга. Так, для птиц видами-инкубаторами послужили, по всей видимости, представители пресмыкающихся. Внешне это могло выглядеть так. Ящерица делает кладку яиц, а из них появляются не ящерки, а шустрые птенчики. Не так уж фантастичен этот сюжет, ибо для его реализации требуется всего лишь переписать программу эмбрионального развития и запустить её вместо обычной. Конечно, если задача ставится шире и включает в себя обеспечение жизнеспособности птенчиков, то в соответствии с новой анатомией требуется переделать физиологию. Следует также предусмотреть хотя бы простейшие инстинкты для выживания – как замирать, чтобы не быть сожранными своими же “родителями”, и что клевать, чтобы не подохнуть с голоду. Все эти новшества в программном обеспечении должны быть разработаны заранее, а их “ввод в действие” должен проводиться на этапе осуществления кодировок у зародыша, т.е. на этапе закрепления соответствий между его генами и теми пакетами программ, ключами к которым эти гены являются.
Теперь обратим внимание на следующее. С одной стороны, и облик, и физиология, и поведение у представителей вида-выводка могут весьма сильно отличаться от таковых у представителей вида-инкубатора. С другой стороны, геномы первого поколения вида-выводка не должны выходить за рамки генофонда вида-инкубатора, чтобы зачатие и развитие эмбрионов не осложнялись проблемами генетической несовместимости. Вот так задачка: геном дитяти должен состоять, как обычно, лишь из генов папаши и мамаши, а генотип дитяти должен включать в себя совершенно несвойственные папаше и мамаше новшества! Но эта, казалось бы, неразрешимая задачка решена Программистами до смешного просто. Вот в чём секрет. У самой разнообразной живности значительные части содержимого хромосом “ни за что не отвечают”, поскольку ничего не кодируют. Генетики их так и называют: некодирующие участки ДНК. Назначение этих некодирующих участков до сих пор остаётся загадкой для науки. Пользы по жизни от них никакой, и неясно, для чего из поколения в поколение перетаскиваются эти многочисленные “генетические пустышки”, обзываемые некоторыми американскими специалистами “генетическим мусором”. Напротив, мы полагаем, что некодирующие участки ДНК – это стратегические запасы, которые нужны, главным образом, для того, чтобы при необходимости можно было использовать любую живность, имеющую их, в качестве вида-инкубатора. Действительно, именно из некодирующих участков ДНК родителей формируют гены, которыми кодируют все новые свойства дитяти. И наоборот, родительские свойства, которые для дитяти ни к чему, отключают, переводя соответствующие участки ДНК в разряд некодирующих. Как можно видеть, в результате такого вмешательства кодирующая часть генома дитяти может значительно отличаться от того её варианта, который бы получился при обычном, автоматическом режиме кодировок, когда она состояла бы лишь из кусочков кодирующих частей геномов родителей. Но “ручная” переделка кодирующей части генома дитяти не изменяет его геном в целом, поэтому проблем генетической несовместимости не возникает.
Кстати, начиная со второго поколения вида-выводка, возможна постепенная доработка его генофонда, чтобы в итоге он всё-таки отличался от генофонда вида-инкубатора. Впрочем, известно, что отличия между генофондами даже на несколько процентов обычно достаточно для того, чтобы соответствующие виды были “генетически обособлены” друг от друга. Ведь при небольшом различии генофондов в целом, различия между их кодирующими частями могут быть существенными! Теперь становится понятно, почему на уровне составов геномов не обнаруживается восходящих эволюционных линий; например, почему геном Homo sapiens так мало отличается от геномов мыши или свиньи.
Как мы постарались показать, успешное созидание нового биологического вида – это филигранная операция, проводимая с программного уровня реальности. Переделка программного обеспечения, перекодировки – для этого требуются соответствующая квалификация и знание кодов доступа. Смешно даже пытаться получить новый биологический вид, не умея работать на программном уровне, а используя методы искусственной селекции или генной инженерии.
Появление новых биологических видов в результате работы Программистов – эта версия прекрасно согласуется с накопившимися фактами, которые не укладываются в теорию естественной эволюции. Особо отметим, что вышеизложенная модель легко объясняет феномен внезапного появления нового вида безо всяких переходных форм. Умея осуществлять такого рода феномены, Программисты имели возможность провернуть всю биологическую эволюцию за чрезвычайно короткие сроки по сравнению с теми, которые требуются теоретикам естественной эволюции. Например, если допустить, что в среднем на восходящих эволюционных ветвях млекопитающих, начиная от первых их представителей, новый вид выводился по 10 раз, да через 10 поколений, да смена поколений происходила через 20 лет, то вся “эволюция” млекопитающих могла уложиться всего в две тысячи лет!
Сейчас мы, по-видимому, переживаем эпоху, когда созидание новых биологических видов приостановлено. Никто отчего-то не дивергирует; действует лишь естественный отбор, едва успевающий устранять нарушения равновесий, которые устраивают в природе неразумные люди. А рабочий метод естественного отбора всегда один и тот же. Результаты известны – видовое многообразие планеты стремительно сокращается. Но нельзя сказать, что биологическая эволюция закончена. Не будем забывать про завалы некодирующих участков ДНК. Они ещё могут пойти нарасхват! Не нужно нам лишних ферментов!
Специалистам слишком хорошо известно о принципиальном различии между молекулами неживой и живой природы. Косное вещество подвержено действию лишь физических и химических законов. Его молекулы образуются с выделением энергии, а для их разрушения требуется энергию затратить. Высокомолекулярные же соединения в живых организмах, наоборот, образуются с энергозатратами, а при их распаде энергия выделяется. Само существование таких соединений противоречит термодинамике, что в своё время было темой оживлённой дискуссии с участием академиков. Если не рассматривать биомолекулы как непостижимое чудо, то напрашивается вывод: они подвержены не только действию физических и химических законов, но и некоторому дополнительному управлению. А такой вывод делает очевидной ограниченность материалистической доктрины.
Посему школьникам ничего не говорят о парадоксальных свойствах биомолекул, и тем более ничего не говорят о том, что биомолекулы могут иметь какое-то “дополнительное управление”. Вот как рассказывают в школе про биосинтез белков: “Сначала двойная спираль ДНК расплетается, чтобы на её определённом кусочке могла синтезироваться информационная РНК“. Дети спрашивают: “А почему ДНК начинает расплетаться? ” – “А потому, – поучают их, – что по ней начинает ползти фермент ДНК-полимераза, который и расплетает её”. – “А почему эта поли-мараза начинает ползти? ” Этот детский вопрос приводит школьных учителей в ступор, а академиков в бешенство – не оттого, что обнаруживаются грани их незнания, а оттого, что до этих граней даже дети добираются за два шага.
А что же будет, если дети узнают самую страшную тайну молекулярной биологии? Вот эта тайна. Оказывается, молекулы аминокислот, с огромными трудами синтезированные искусственно или взятые из начавших разлагаться трупов, являются неустойчивыми. Хуже того, если им предоставить возможность самопроизвольно соединяться друг с другом, то они в условиях, подходящих для “существования белковых тел”, не образуют пептидных цепочек, поскольку охотнее соединяются не карбоксильной и аминной группами, а другими радикалами. Это нужно как следует уяснить: молекулы белков в природе самопроизвольно не образуются! Откуда же они взялись на планете? …В условиях, когда не было ещё ни фотосинтеза, ни биосинтеза на основе ДНК! Кстати, молекулы ДНК тоже самопроизвольно не образуются. Они ещё более высокоэнергичны, чем молекулы белков.
Да что там белки или ДНК! Как в принципе образуются и существуют даже простенькие молекулы биоорганики, например, те же аминокислоты, если соединённые в них радикалы находятся в более высоких энергетических состояниях, чем разъединённые? Молекулярные биологи ухватились вот за какую соломинку. Они полагают, что весь этот физико-химический абсурд нисколько не удивителен, если синтез биоорганики происходит при участии специфических биокатализаторов – ферментов. Вот как это описано в новейшей энциклопедии для детей (издательства “Аванта+”): “фермент и субстрат подходят друг к другу, как замок к ключу: распознав ‘свой’ субстрат, фермент подстраивается под его ‘капризы’ и готовит для него наиболее удобное ‘посадочное место’. ‘Посадочные места’ на ферменте устроены очень продуманно. Субстраты оказываются расположены друг к другу именно так, как это требуется для определённой реакции между ними. Затем молекула фермента сталкивает их, и в итоге получается нужный продукт“. И вот это всё, простите, вытворяют молекулы? Ну, тогда, действительно, начхать им на термодинамику. Слишком уж “продуманно” у них всё получается. Кстати, кто это всё так здорово продумал? И, кстати, если биосинтез происходит только под прессингом ферментов, то неужели ещё до синтеза первых “нужных продуктов” кто-то заботливо состряпал необходимые для этого ферменты? Весёленькие же дела творились на планете в эпоху её бурной молодости, когда, согласно научному поверью, жизнь изо всех сил пыталась зародиться.
Ну да ладно, не будем ворошить прошлое. Тут и в настоящем проблем хватает! Загвоздка в том, что полный комплект ферментов, якобы требуемый для синтеза биомолекулы, непомерно велик. Так, в той же энциклопедии для детей читаем: “Специализация ферментов достойна восхищения. Каждый из этапов реакции идёт при участии отдельного фермента. Мало того, если в организме осуществляются прямая и обратная реакции, их тоже ведут – и всегда разными ’маршрутами’ – разные ферменты. Например, белки разрушаются протеазами, а синтезируются уже лигазами. Разделение труда есть и внутри каждой молекулы. Это легко понять, сопоставив по размеру молекулы фермента и субстрата – соотношение между ними может достигать 1000: 1 – 10000: 1“. Тут дети догадываются, что молекулы ферментов тоже ведь нужно каким-то образом синтезировать, а для этого потребуются новые ферменты и так далее. Результирующие соотношения между ферментами и “полезным продуктом” настолько чудовищны, что даже дети понимают их нереальность. Нет особой разницы между тем, содержатся ли в организмах полные наборы ферментов постоянно, или ферменты синтезируются лишь по мере надобности. В обоих случаях организмы состояли бы почти сплошь из ферментов.
И это не всё. При нереально огромном количестве требуемых ферментов им приписывают ещё и фантастически высокую эффективность работы. Они якобы “способны увеличить скорость реакции в сотни тысяч и миллионы (а возможно, даже в триллионы) раз“. Совершенно мистически они должны по очереди оказываться в нужном месте в нужное время, обеспечивая требуемую последовательность присоединения радикалов. Существуют теории элементарного акта ферментативного катализа. Но при типичных скоростях биосинтеза макромолекул эти “акты” должны следовать друг за другом с такой быстротой, которая не укладывается ни в какие рамки химической кинетики.
Между тем, вышеназванные парадоксы устраняются, если допустить-таки, что биоорганика в живых организмах может быть подвержена специфическому управлению, которому не подвержено вещество неживой природы. При таком допущении кардинально упрощаются представления о процессах на молекулярном уровне, происходящих в живых организмах. С программного уровня реальности возможно эффективное воздействие на вещество, чтобы атомы и радикалы соединялись в конфигурации, которые не образуются самопроизвольно. Правда, для каркаса таких конфигураций подходят не любые атомы, а только те, которые легко образуют двойную химическую связь, при этом имея в запасе ещё несколько валентных электронов. Самые лёгкие из атомов, обладающие этими свойствами – это атомы углерода, азота и кислорода. Именно они используются для построения структур биомолекул, “невозможных” по меркам неживой природы.
Ключевую роль в этих структурах играют двойные связи. В “Ступенях испепеляющих” упоминалось о том, что химическая связь представляет собой циклический процесс. Связующий электрон “переключается” из состава одного связуемого атома в состав другого и обратно. При двойной связи два таких процесса происходят параллельно. Кроме того, при двойной связи возможно явление, о котором говорил Полинг, а именно, циклические переходы связующих электронов с одной связочки на другую. В данном случае такие переходы возможны, если они происходят синхронно. Причём, если равны обе частоты “переключений”, то максимальная частота синхронных переходов может быть равна частоте “переключений”. Тогда результирующая связка будет иметь двойную синхронизацию. Другими словами, она будет являться дважды резонансной. Как нам представляется, такие дважды резонансные связки как раз и обеспечивают высокоэнергичные соединения атомов, которые характерны для биомолекул.
Самопроизвольное образование дважды резонансных связок крайне маловероятно. Валентные электроны в атомах находятся в различных энергетических состояниях, и, если двойная связь образуется самопроизвольно, то частоты “переключений” связующих электронов, как правило, различаются. Поэтому такая связь имеет, как правило, нерезонансный характер. Но примечательно, что частоты “переключений” и синхронных переходов зависят от компоновки атомарных частотных гнёзд, в которых удерживаются валентные электроны. Значит, частотами “переключений” и синхронных переходов можно управлять, изменяя компоновку частотных гнёзд в связуемых атомах.
И что же мешает этому? Если атомарные структуры держатся не благодаря электромагнитному взаимодействию, а благодаря особенностям Физической Арены, формируемым с программного уровня, т.е. если компоновка частотных гнёзд в атоме диктуется программными средствами, то с помощью дополнительной программы можно изменять эту компоновку. Таким образом, воздействиями с программного уровня, через создание благоприятных условий для образования дважды резонансных связок, возможно выстраивать молекулярные структуры, которые не образуются из “неуправляемых” атомов. Следует подчеркнуть, что эти воздействия с программного уровня являются не энергетическими. Они лишь перераспределяют энергии на физическом уровне, так что закон сохранения физической энергии не нарушается.
Вспомним: гипотеза о том, что в биосинтезе непременно участвуют ферменты, появилась как попытка объяснения биохимических процессов, “невозможных” по меркам неживой природы. Но если вещество живой природы охвачено эффективным управлением с программного уровня, то проблема разрешается гораздо проще. Не требуются никакие катализаторы для тех биохимических реакций, которые и без катализаторов полностью управляемы. Это, конечно, не означает, что ферменты в организмах не нужны вовсе. Они нужны, но для работы с поступающими в организмы чужеродными биомолекулами, к которым не подключено “своё” управление. Аккуратно распиливать чужие биомолекулы на кусочки – вот работа для ферментов, например, для пищеварительных. Вклиниться в “переключения” химической связи и разорвать её возможно, если у фермента частота смен электронных конфигураций в радикале, работающем в качестве молекулярной пилы, превышает частоту смен электронных конфигураций в распиливаемой связи. Не всегда это так, и у молекулярной пилы может “не хватать оборотов” для того, чтобы справиться с высокоэнергичной связкой. Но ведь молекулярная пила своего фермента управляема, и можно добавить ей оборотов! Здесь – разгадка того феномена, что химически один и тот же фермент при одних и тех же физических условиях либо распиливает мишень, либо нет. Наука комментирует это так, что мало иметь фермент – он работает, лишь будучи активирован!
Итак, активированные ферменты хороши для того, чтобы с их помощью ломать чужие биомолекулы. Строить же свои биомолекулы – это совсем другая задача. Здесь ферменты не требуются. Обрисуем вкратце возможности управляемого биосинтеза с применением вышеописанных воздействий на вещество.
Управлять атомами можно было так, чтобы первые биомолекулы собирались, что называется, на пустом месте в каком-нибудь проницаемом для солнечного света растворе. В энергию резонансных связей можно было превращать энергию возбуждённых электронных состояний, которые образуются в атомах на свету. Этот принцип используется при фотосинтезе. Всё бы хорошо, но скорость управляемого биосинтеза “на пустом месте” ограничена тем, что при заранее известном дизайне всей молекулы возможно лишь последовательно присоединять очередные атомы или радикалы к уже синтезированному кусочку. Казалось бы, по-другому и быть не может. Но с давних пор применяется гораздо более эффективный способ управляемого биосинтеза: с использованием молекул-матриц, обеспечивающих возможность многоканальной параллельной сборки нужной молекулы!
Молекула-матрица всего лишь задаёт взаиморасположение компонентов синтезируемой молекулы. Управление атомами для образования высокоэнергичной связи включается тогда, когда подходящий атом или радикал оказывается в подходящем объёмчике, т.е. “на своём месте” в составе будущей молекулы. Каждый такой объёмчик, в котором может включиться это самое управление, должен иметь пространственную привязку. Роль вещественных маркеров для этой привязки могут играть, например, периферийные атомы на недостроенной молекуле. Но при этом, как уже отмечалось, биосинтез возможен лишь путём последовательной достройки. Зато молекула-матрица – это полный набор вещественных маркеров, задающих строение нужной молекулы.
Управляемый матричный биосинтез может выглядеть не так уж сложно, как это обычно полагают. В самых общих чертах, может происходить следующее. Молекула ДНК спроектирована восхитительно: одномоментным воздействием с программного уровня возможно так ослабить связи на её заданном участке, что соответствующий отрезок одной из цепочек вывалится из неё с сохранением последовательности нуклеотидов. Через некоторое время – еще одно переключение с программного уровня, и брешь в цепочке быстро заполняется по принципу комплементарности. Таким образом, ДНК восстанавливается в исходное состояние, а извлечённый из неё нужный отрезок цепочки нуклеотидов, который учёные в шутку называют информационной РНК, используется как молекула-матрица. Важно отметить, что соответствия между триплетами нуклеотидов в молекуле-матрице и аминокислотами в синтезируемой молекуле белка заданы отнюдь не на основе химического сродства между триплетами и аминокислотами. Эти соответствия заданы программными средствами, о чём свидетельствует хотя бы тот факт, что одна и та же аминокислота может кодироваться различными триплетами. Итак, щелчок с программного уровня – и начинается строительство. У молекул аминокислот, которые попадают на “свои” места, пространственно обозначенные с помощью молекулы-матрицы, включается механизм образования пептидных связей с соседями. Это чудесное действо происходит сразу вдоль всей цепочки “посадочных мест”, обозначенных в пространстве молекулой-матрицей. При этом исключена возможность ошибочной посадки аминокислоты из-за неоднозначного прочтения цепочки триплетов, ведь чтение можно начать с любого места. Такие ошибки были бы возможны, если посадочные места аминокислот были бы обусловлены химической согласованностью с триплетами. Иное дело – посадочные места, формируемые программными средствами. Положения этих посадочных мест относительно молекулы-матрицы заданы однозначно. И вот, поскольку здесь синтез ведётся сразу на всей длине молекулы, то скорость его несоизмеримо выше, чем при последовательном достраивании. Матричный биосинтез даёт возможность штамповать первичные структуры белков, тратя на гигантскую молекулу ненамного больше времени, чем на самую простенькую. Похоже, матричный биосинтез выжимает из химической кинетики атомов и радикалов всё, что только можно. Если, конечно, ферменты под ногами не путаются. Это не шутка: если хотите замедлить матричный биосинтез “в триллионы раз”, то окружите молекулу-матрицу толпой малоподвижных макромолекул, и дело будет в шляпе.
Молекулярные биологи вольны не верить в то, что химические процессы в живых организмах управляются с программного уровня, и вольны не верить в то, что кто-то организовал это управление. Пусть эти неверующие молекулярные биологи продолжают свои игры в ферментюльки. Как резонируют белк и .
Молекулярная биология полагает, что последовательность аминокислот в пептидной цепочке полностью определяет строение молекулы белка, а значит, и её физико-химические свойства, а значит, и её биологические функции. Из этого подхода следует, что если искусственно воспроизвести такую же последовательность аминокислот, как и в каком-либо “природном” белке, то искусственный белок будет представлять собой в биохимическом отношении то же самое, что и “природный”. Опыт опровергает эти наивные надежды. Искусственные белки, в лучшем случае, не выполняют никаких биологических функций. Хуже того – они оказываются сильными аллергенами.
Логично предположить, что “визитная карточка” белка представлена не только последовательностью аминокислот, но и некоторым дополнительным признаком. И, если этот дополнительный признак несут не аминокислоты, как таковые, то следует поискать его в пептидных связях, которыми аминокислоты сцеплены друг с другом. К счастью, не нужно далеко ходить. На сегодня можно считать уже доказанным, что пептидные связи являются резонирующими, т.е. что в них происходят циклические переключения конфигураций химических связей. Так, читаем (Комов и Шведова, ”Биохимия”, 2004): “Пептидная связь не является полностью одинарной. Её длина составляет 0.132 нм, что является средним значением между истинной одинарной связью C-N (0.149 нм) и истинной двойной связью C=N (0.127 нм). По некоторым данным, пептидная связь является частично двойной и частично одинарной. Обе структуры динамичны, и между ними имеются взаимные переходы“. Но в том же учебнике приведена химическая схема пептидного соединения двух аминокислот, которое прямо-таки “железобетонное”. Не просматривается даже намёка на “взаимные переходы”. Начинают во здравие, кончают за упокой!
Давайте же будем последовательны! Схему химических связей в аминокислоте обычно представляют следующим образом:
где R - характерный радикал. Пептидное соединение образуется при сцепке карбоксильной группы одной аминокислоты с аминной группой другой. При этом из карбоксильной группы “изымается” гидроксильная группа ОН, а из аминной – атом водорода Н. В результате, как нам представляется, образуется резонирующее соединение, две конфигурации которого, попеременно сменяющие друг друга, изображены ниже.
Переключения между такими конфигурациями предполагают завидную синхронность, т.к. переходы химических связей затрагивают шесть различных атомов. Но мы не усматриваем в этом большой проблемы, поскольку допускаем, что управление переключениями электронных конфигураций возможно с программного уровня реальности (см. “Не нужно нам лишних ферментов! “). При этом физико-химические законы ничуть не нарушаются. Свойства атомов остаются такими же, как и при отсутствии управляемых переключений: при обеих конфигурациях водород образует одну связь, кислород – две, азот – три, а углерод – четыре. И, конечно, расстояние между атомами углерод-азотной пары в пептидной связи должно иметь значение, промежуточное между значениями для случаев чисто одинарной и чисто двойной связей, поскольку при конфигурации 1 связь в этой углерод-азотной паре двойная, а при конфигурации 2 – одинарная.
Только что обрисованная картина резонанса в пептидном соединении позволяет сделать некоторые интересные выводы. Ведь, если переключения электронных конфигураций в пептидном соединении производятся благодаря управлению с программного уровня, то частота этих переключений является управляемым параметром! Отсюда сразу же просматривается то, что же именно дополняет последовательность аминокислот в “визитной карточке” белка: это специфические частоты резонансов в пептидных соединениях тех или иных пар аминокислот. Тогда “природные” белки имеют поразительную защиту от подделок. Синтезируя белок физико-химическими методами, возможно подделать последовательность аминокислот, но невозможно подделать частоты пептидных резонансов.
Дополнительная “степень защиты” белков, о которой идёт речь, имеет большой биологический смысл. Прежде всего, эта “степень защиты” повышает эффективность и надёжность генетической обособленности. У генетически обособленных организмов могут различаться частоты пептидных резонансов между одними и теми же парами аминокислот, и, соответственно, могут различаться полные наборы частот пептидных резонансов. “Своим” будет считаться тот белок, у которого правильны как последовательность аминокислот, так и частоты соответствующих пептидных резонансов.
Насколько же это облегчает работу иммунной системы по распознанию “чужих” белков! Когда пептидная цепочка свёрнута в глобулу, то иммуноглобулины могут контактировать лишь с некоторыми участками этой цепочки, которые расположены на периферии глобулы. При этом большая часть первичной структуры цепочки остаётся недоступной, и признак правильности частот пептидных резонансов выходит по значимости на первое место. Не будь этого признака, нам пришлось бы допустить, что для распознания “чужих” глобул иммуноглобулины используют какое-то неизвестное науке чутьё.
Впрочем, главное, что даёт характеристический набор частот пептидных резонансов - это обеспечение дополнительной специфичности ”своих” белков. Эта специфичность требуется для управления белками в режиме селективного доступа, по следующим принципам. Во-первых, “я” управляю только ”своими” белками и никакими больше. Во-вторых, ”своими” белками управляю только “я” и никто больше.
Тут биохимики, конечно, вклинятся с заявлением о том, что никакого управления белками нет; что, действительно, белки вытворяют много премилых вещей, но всё это происходит абсолютно самопроизвольно. Свежо предание! Начать с того, что абсолютно самопроизвольно происходит синтез биополимеров, которые не встречаются в неживой природе. Делать им там нечего, а вот в живых организмах у них множество функций, оттого они там самопроизвольно и синтезируются. Что интересно: после своего самопроизвольного синтеза они самопроизвольно формируют себе вторичную структуру, скрепляясь водородными связями, а затем и третичную, скрепляясь связями дисульфидными. У белка средней длины имеется огромное число устойчивых конфигураций вторичной и третичной структур, но абсолютно самопроизвольно формируются именно те, которые как раз требуются для решения той или иной неотложной задачи. Само собой, решение задачи тоже происходит абсолютно самопроизвольно, как будто её никто и не ставил. Ну, это понятно - белки-то синтезировались именно такие, какие и требовались. Кстати, задачи, которые ”никто не ставил”, бывают такими нетривиальными, что молекуле белка для их решения приходится абсолютно самопроизвольно перестраивать свою конфигурацию, чтобы подобрать оптимальную. Причём, те локальные задачки, которые решают отдельные молекулы белка, оказываются - опять же, абсолютно самопроизвольно - изумительно согласованы друг с другом, так что в результате решается более глобальная задачка, благодаря чему продолжает трепыхаться жизнь в организме. И так далее, и так далее. Ибо если считать, что всё это происходит абсолютно самопроизвольно, то уж нужно переть до конца. И в конце сделать логичный вывод о том, что взлёты теоретической мысли биохимиков, которые они расценивают как свои достижения, не более чем абсолютный самопроизвол.
У нас же подход иной. ”Трепыхание жизни” в организме поддерживается благодаря работе мощного пакета программ, в котором предписано, что требуется выполнить с помощью молекул белков в той или иной ситуации, как синтезировать подходящие для этих целей белки, и как затем управлять этими белками, чтобы они выполняли задуманное. Если судить по внешним проявлениям, то компоновка этого мощного пакета программ, его иерархия, ветвление, селективный доступ - всё это поразительно напоминает аналогичные атрибуты программ, которые сегодня пишутся на языках программирования высокого уровня. Сегодня, серьёзно занимаясь программированием, легко постигнуть главную тайну ”существования белковых тел”. Кто-то разрабатывал программы, благодаря которым это существование влачится. Мы называем этих разработчиков Программистами.
О том, каким образом управление с программного уровня проводится на физический уровень, т.е. на вещество, мы уже говорили в предыдущих работах. Ключевую роль при этом проведении управления играет обеспечение его селективности: управлять желательно только ”своим” веществом, но не ”чужим”. И здесь как нельзя кстати оказывается такой дополнительный идентификационный признак у молекул белков, как индивидуальный набор частот пептидных резонансов. Программное управление подключается только к тем пептидным цепочкам, которые резонируют ”правильно”. |
Последнее изменение этой страницы: 2017-05-11; Просмотров: 188; Нарушение авторского права страницы