|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Вопрос 5.Формула и диаграмма цветка: понятие и правила написания.
Начиная с XIX в. в учебной работе и в научных исследованиях для большей наглядности используют формулы и диаграммы. Формула цветка - условное обозначение его строения с помощью букв латинского алфавита, символов и цифр. При составлении формулы употребляют следующие обозначения: Р - простой околоцветник (perianthiutn); Ca (или К) - чашечка (calyx); Со (или С) - венчик (corolla); A - андроцей (androeceum); G - гинецей (gynoeceum); знак *, помещаемый в начале формулы, указывает на актиноморфность цветка; стрелка вверх - на его зигоморфность. Обоеполый цветок, обозначается знаком, обозначенным на рис. 222, А; мужской - на рис. 222, Б; женский - на рис. 222, В. Знак " +" указывает на расположение частей цветка в двух или нескольких кругах либо на то, что части, разделенные этим знаком, противостоят друг другу. Скобки означают срастание частей цветка. Цифра рядом с символом указывает на количество членов данной части цветка; черта под цифрой, обозначающей число плодолистиков в гинецее, свидетельствует о том, что завязь верхняя; черта над цифрой - завязь нижняя; черта от цифры - полунижняя завязь. Большое и неопределенное число членов обозначается знаком бесконечность. Диаграмма цветка более наглядна, чем формула. Она представляет условную схематическую проекцию частей цветка на плоскость и отражает их число, относительные размеры и взаимное расположение, а также наличие срастаний ( рис. 99 ). Кроме того, на диаграмме нередко указывается расположение кроющего (прицветного) листа, прицветничков и оси соцветия или побега, несущего цветок. Прицветник, прицветнички и чашелистики чаще изображаются скобками с килем (фигурными скобками) различного размера, лепестки - круглыми скобками, тычинки в виде среза через пыльник или для упрощения в виде затушеванного эллипса, гинецей - также в виде среза через завязь или завязи (для апокарпного гинецея ) с прорисовкой места плацентации и семязачатков, через которые прошел срез. Вопрос 6. Способы и формы опыления. Приспособления растений к перекрестному опылению. Имеется два основных типа опыления: самоопыление (когда растение опыляется собственной пыльцой) и перекрёстное опыление. При перекрёстном опылении растения могут давать два основных типа растений: однодомные и двудомные. Перекрёстное опыление требует участия посредника, который бы доставил пыльцевые зёрна от тычинки к рыльцу пестика; в зависимости от этого различают следующие типы опыления: · Биотическое опыление (при помощи живых организмов) · Энтомофилия — опыление насекомыми Пыльца цветков, как правило, крупная и очень клейкая. Некоторые виды растений (например, лютики) имеют чашевидную форму цветка, чтобы залезшее в него насекомое «пачкалось» о пыльцу, улучшая процесс опыления. · Зоофилия — опыление при помощи позвоночных животных: птицами Искусственное опыление — перенесение пыльцы с тычинок на пестики цветков при посредстве человека[2]. · Абиотическое опыление · Анемофилия — опыление с помощью ветра, очень распространено у злаков, большинства хвойных и многих лиственных деревьев. · Гидрофилия — опыление при помощи воды, распространено у водных растений. Около 80, 4 % всех видов растений имеют биотический тип опыления, 19, 6 % опыляются при помощи ветра. Гейтоногамия — соседнее опыление, опыление рыльца пестика одного цветка пыльцой другого цветка того же растения[3].
На протяжении исторического развития ентомофилии у растений выработались различные приспособления, препятствующие самоопылению и способствующие попаданию на рыльца пыльцы с других цветков. Они обусловлены строением и физиологическими особенностями цветков. К таким приспособлениям относятся: размещение отдельно мужских и женских половых органов, то есть однополость цветков у однодомных (огурец, тыква) и двудомных растений (ива, конопля); гетеростилия или разностолбчатость, когда у одних цветков высокие столбики пестиков и короткие тычинки, а у других - низкие столбики и длинные тычинки, например, у гречихи, плакуна иволистного; разновременное созревание тычинок и пестиков, вследствие чего рассыпание пыльцевых зерен из пыльников не приводит к опылению из-за незрелости рылец (у подсолнечника и других растений семейства сложноцветных), или окончание функционирования рылец до созревания собственной пыльцы (у яблони, груши); самостерильность, характеризующаяся неспособностью пыльцы прорастать на рыльце пестика или несовместимостью половых клеток при оплодотворении, вследствие чего наступает самонеплодность, например, у плодовых деревьев, когда опыление цветков даже в пределах одного сорта, не дает урожая. Для привлечения пчел, шмелей и других полезных насекомых, которые осуществляют перекрестное опыление, у растений возникли различные приспособления: выделение цветами ароматических веществ, окраска лепестков и других частей в разные цвета, формирование хорошо заметных крупных соцветий, своеобразное строение цветков. Но самым сильным фактором привлечения при разной интенсивности посещения цветков и переноса пыльцы есть пища для насекомых - нектар и пыльца. Собирая пищу, пчелы вошли в тесную связь с растениями и приносят им огромную пользу. Неопыленные цветки отмирают плодов и семян не образуют.
№7 Андроцей, типы андроцеев, строение пыльника. Микроспорогинез развитие и строение мужского гаметофита. Андроцей (от греч. aner, род. падеж andró s — мужчина и oikiа — жилище), совокупность всех мужских органов цветка — тычинок. Андроцей считается мужским органом полового размножения. Типы андроцея: · 1 - четырехсильный (у крестоцветных ), · 2 - двусильный (характерен для многих губоцветных ), · 3 - двубратственный ( бобовые подсемейства мотыльковых ), · 4 - андроцей со склеенными в трубку пыльниками ( сложноцветные ). По длине тычиночных нитей: · двухсильный андроцей – две тычинки длиннее остальных, · четырехсильный – четыре тычинки длиннее остальных и т.п. Строение тычинок: У тычинки можно различить нитевидную часть – тычиночную нить и расширенную часть – пыльники. Пыльник состоит из двух тек (сумок), соединенных связником. Каждая тека содержит два пыльцевых гнезда. В центре гнезда располагаются археспориальные клетки, из которых в дальнейшем в результате мейоза образуются микроспоры, прорастающие прямо в пыльниках в мужской гаметофит – пыльцевое зерно. Стенка пыльника имеет сложное строение. Снаружи располагается эпидерма, затем эндотеций, средний слой из паренхимных клеток, самый внутренний слой – тапетум. Клетки тапетума и среднего слоя имеют густую цитоплазму и служат для питания развивающейся пыльцы. Клетки эндотеция имеют утолщенные фиброзные пояски. После созревания пыльцы клетки эндотеция постепенно отмирают, усыхают и сильно сокращаются, пыльники вскрываются по этим пояскам. Вскрытие пыльника происходит под давлением, что способствует разбрасыванию пыльцевых зерен.У некоторых видов развиваются стерильные тычинки, без пыльников. Такие тычинки называют стаминодиями. Они иногда становятся нектарниками. Микроспорогенез – это процесс образования микроспор в микроспорангиях, которыми являются гнезда пыльника. Клетки спорогенной ткани несколько раз последовательно делятся митозом. В результате этого формируются диплоидные (2n) микроспороциты (материнские клетки микроспор). Каждый микроспороцит делится однократно мейозом, в результате чего возникает тетрада (четыре) гаплоидных (n) микроспор. Микроспора – это тонкостенная клетка с одним гаплоидным ядром. Чаще всего стадия тетрады кратковременна, поэтому микроспоры быстро обособляются друг от друга. Иногда тетрады микроспор сохраняются, образуя в последующем тетрады пыльцевых зерен (рогоз, элодея, вересковые, росянка). Все развитие мужского гаметофита, включая образование мужских гамет, сводится лишь к двум митотическим делениям. Первое из этих делений происходит всегда под защитой оболочки микроспоры, которая превращается в новое образование — пыльцевое зерно. Второе деление совершается в одних случаях в пыльцевом зерне, а в других — лишь позднее, в пыльцевой трубке. В результате зрелые пыльцевые зерна бывают двух клеточными или трехклеточными, причем двухклеточные встречаются значительно чаще, чем трехклеточные, и известны приблизительно у 70% цветковых растений. Еще в 1926 г. известный немецкий эмбриолог растений П. Шюргоф пришел к выводу, что в процессе эволюции трех клеточная пыльца возникла из двухклеточной. Он справедливо считал, что прохождение обоих делений внутри пыльцевого зерна, в то время как последнее еще не отделилось от материнского растения, является прогрессивным, так как в этом случае развитие мужского гаметофита происходит под надежной защитой спородермы и более быстро. Действительно, двухклеточная пыльца характерна для многих относительно примитивных групп, в том числе для всех представителей порядков магнолиевых, лавровых, нимфейных, лютиковых, а трехклеточная пыльца встречается чаще у более подвинутых порядков, в том числе у всех сложноцветных и злаков. Перед началом первого митотического деления ядро микроспоры переходит из центрального положения в периферическое. В результате деления образуются две, как правило, неравные клетки — маленькая, обычно пристенная генеративная клетка и большая сильно вакуолизированная клетка-трубка (широко известная также под неудачным названием вегетативной клетки). Генеративная клетка вначале прилегает к оболочке микроспоры (чаще всего против апертуры), но вскоре становится выпуклой, все более и более вдается в клетку-трубку, наконец, совсем отделяется от оболочки и полностью погружается в цитоплазму клетки-трубки, где лежит свободно. Генеративная клетка окружена топкой, прозрачной и легкопроницаемой оболочкой и принимает эллипсоидальную или веретенообразную форму. Создается совершенно уникальная ситуация, когда одна клетка помещается внутри другой. Дальнейшая судьба этих двух клеток глубоко различна. В результате вторичного деления (происходящего внутри оболочки пыльцевого зерна или позднее в пыльцевой трубке) из генеративной клетки образуются две безжгутиковые мужские гаметы — спермии, а клетка-трубка больше не делится и дает начало пыльцевой трубке. Таким образом, мужской гаметофит цветковых растений достиг наивысшей степени упрощения: он совершенно лишен как проталлиальных клеток, так и антеридия и состоит всего лишь из двух клеток, одна из которых, делясь, образует две очень упрощенные гаметы.
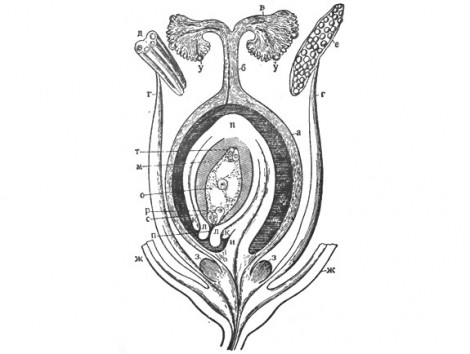
№8 строение семяпочки. Мегаспорогинез. развитие и строение женского гаметофит а. Место прикрепления семязачатка ( семяпочки ) к семяножке называется рубчиком. Нижняя часть семяпочки, от которой отходят интегументы, или покровы семяпочки, называется халазой (рис. 163). В завязи синкарпного гинецея плодолистики могут срастаться только краями, по периферии, образуя тогда одногнездную завязь; или же плодолистики, заворачиваясь, срастаются краями внутри завязи, разделяя ее на ряд гнезд ( многогнездная завязь ). Зрелая семяпочка снаружи покрыта интегументом — одним, или чаще двумя (наружным и внутренним). Интегументы на верхнем конце не срастаются, образуя пыльцевход ( микропиле ). Далее внутрь располагается нуцеллус (ядро семяпочки). Сильно развитый нуцеллус с двумя многослойными интегументами имеется у проще организованных раздельнолепестных растений. Сложнее организованным спайнолепестным (за рядом исключений) свойствен небольшой нуцеллус с одним (внешним) интегументом. Внутренний интегумент у них не развивается. Таким образом, один покров спайнолепестных не гомологичен одному интегументу голосеменных. В верхней части нуцеллуса расположен зародышевый мешок. Зародышевый мешок в верхней, обращенной к микропиле части содержит яйцевой аппарат, состоящий из голых или одетых белковой оболочкой клеток. Одна из них, центральная, более крупная — яйцеклетка, две боковые — так называемые синергиды (клетки-помощницы). На противоположном конце зародышевого мешка имеются три клетки, называемые антиподиальными ( антиподы ). В центральной части зародышевого мешка расположено вторичное ядро зародышевого мешка, образовавшееся от слияния двух полярных ядер. В таком виде зародышевый мешок готов к оплодотворению.
Мегаспорогенез – это процесс формирования мегаспор в нуцеллусе семязачатка.В нуцеллусе вблизи микропиле обособляется, увеличивается одна (реже несколько) клета археспория. Эта клетка становится материнской клеткой мегаспор, она имеет диплоидный набор хромосом (2n). Материнская клетка однократно делится мейозом, в результате возникает тетрада гаплоидных мегаспор (четыре по 1n). Мегаспоры располагаются линейно от микропиле к халазе У большинства цветковых растений (вероятно, не менее чем у 80%) начало женскому гаметофиту дает одна из четырех потенциальных мегаспор тетрады путем трех последовательных митотических делений. Этот тип образования женского гаметофита называется моноспорическим. Его развитие происходит следующим образом ( рис. 28Б ). В результате первого деления ядра функционирующей мегаспоры образуются два ядра, которые расходятся к полюсам (микропилярному и халазальному) сильно удлиняющейся мегаспоры, а между ними образуется крупная вакуоль. Далее каждое из этих двух ядер делится синхронно еще два раза и в результате у каждого полюса образуются по четыре ядра. Это восьмиядерная стадия развития женского гаметофита. От каждой из двух полярных четверок входит по одному ядру в центральную часть зародышевого мешка. Эти так называемые полярные ядра сближаются в центральной части и сливаются или сразу же, или только перед оплодотворением, а иногда и вовсе не сливаются. В результате слияния полярных ядер образуется ядро центральной клетки женского гаметофита, называемое часто вторичным ядром зародышевого мешка. Основной особенностью моноспорического типа образования женского гаметофита является последовательное заложение клеточных перегородок при мейотических делениях и образование обособленных мегаспор. Совершенно иной характер имеет мегаспорогенез, предшествующий образованию биспорического (двухспорового) и тетраспорического (четырехспорового) гаметофитов. Изменения в мегаспорогенезе заключаются в подавлении заложения клеточной перегородки при мейотическом делении, вследствие чего вместо тетрад мегаспор возникают неклеточные образования, называемые ценоцитами (от греч. koinos — общий и kytos — сосуд, клетка), которые и развиваются далее в женские гаметофиты. №9 Типы гинецеев и завязи, признаки определения числа плодолистиков. По количеству пестиков гинецей делят: 1) на простой – состоит из одного пестика (бобовые, капустные, астровые, мятликовые, махорка, мак) 2) сложный – состоит из двух или нескольких пестиков (сусак, земляника, лютик, шиповник). По числу плодолистиков и способу их срастания различают следующие типы гинецея 1) монокарпный – состоит из одного плодолистика. Края единственного плодолистика сходятся и срастаются, на месте их срастания образуется брюшной шов. В результате формируется одногнездная завязь (большинство бобовых, слива, вишня, черемуха); 2) апокарпный – состоит из нескольких свободных (несросшихся) плодолистиков, каждый из которых образует свой пестик (лютик, земляника, малина, шиповник); 3) ценокарпный – состоит из одного пестика, который образован при срастании нескольких плодолистиков. В зависимости от способа срастания плодолистиков и числа гнезд завязи различают три типа ценокарпного гинецея: – синкарпный – края плодолистиков заворачиваются внутрь, срастаются своими боковыми поверхностями и образуют одну завязь, разделенную на гнезда, – число гнезд равно числу плодолистиков (тюльпан, картофель, яблоня); синкарпный гинецей происходит из апокарпного в результате бокового срастания апокарпных плодолистиков, для этого типа характерна центрально-угловая плацентация; – лизикарпный – формируется из синкарпного гинецея путем растворения боковых стенок внутри завязи, поэтому образуется одногнездная завязь, в центре которой сохраняется ось, или колонка (гвоздичные, первоцветные), т. е. плацентация колончатая; – паракарпный – формируется из синкарпного гинецея путем срастания только краев соседних плодолистиков, в результате образуется одногнездная завязь с постенной плацентацией (тыквенные). По типу расположения в цветке завязи называются: Верхняя (свободная) завязь - прикрепляется основанием к цветоложу, не срастаясь ни с какими частями цветка (в этом случае цветок называют подпестичным или околопестичным). Нижняя завязь - находится под цветоложем, остальные части цветка прикрепляются у её вершины (в этом случае цветок называют надпестичным). Полунижняя завязь - срастается с другими частями цветка, гипантием или цветоложем, но не у самого верха, верхушка её остается свободной (в этом случае цветок называют полунадпестичным).
№10 Соцветия: строение, классификация, биологическое значение Соцветие - это побег или система побегов, несущих цветки. На узлах осей соцветия располагаются такие же листья, как на вегетативной части побега, или видоизмененные, утратившие способность к фотосинтезу, - прицветники, а на узлах цветоножки - прицветнички. . Различают два типа соцветий: сложные, когда цветки располагаются на разветвлениях главной оси, и простые, когда цветки с цветоножками или без них располагаются непосредственно на главной оси.
Сложные соцветия в зависимости от способа нарастания осей делят на: симподиальные (определенные) - ось заканчивается цветком, распускание цветков идет от верхушки к боковым ветвям или центробежно, если цветки расположены в одной плоскости; и моноподиальные (неопределенные) - ось нарастает неопределенно долго, расцветание цветков идет от основания к верхушке или центростремительно, если цветки расположены в одной плоскости. Сложные симподиальные соцветия: · монохазий - главная ось заканчивается цветком; под ним образуется ось второго порядка, также увенчанная цветком, и т. д.; если подцветочные оси отходят в одну сторону, то образуется завиток, если же попеременно то в одну, то в другую сторону - образуется извилина; завиток, у которого боковые разветвления укорочены, называют клубочком; · дихазий - под цветком образуются две супротивные оси, каждая из которых также заканчивается цветком и также дает две подцветочные оси, повторяющие такой же способ ветвления, и т. д.; · плейохазий - от главной оси, несущей один верхушечный цветок, отходит больше двух подцветочных осей, образующих мутовку из монохазиев или дихазиев; · тирс - на главной оси располагаются указанные выше соцветия, чаще имеет пирамидальную форму. Сложные моноподиальные соцветия: · метелка - очень разветвленное соцветие, нижние боковые разветвления ветвятся сильнее, чем верхние; · щиток - метелка, у которой цветки расположены более или менее в одной плоскости; · сложный колос - главная ось образует разветвления, на которых расположены цветки без цветоножек; эти разветвления называют колосками; · сложный зонтик - расстояния между осями второго порядка укорочены, и они отходят от верхушки оси первого порядка; расстояния между цветоножками укорочены, и они расположены на верхушке оси второго порядка; нередко листья у основания осей второго порядка образуют общую обвертку, а у основания цветоножек - частную обвертку. Кроме перечисленных имеются сложные соцветия, называемые агрегатными. Они образованы сочетанием различных типов соцветий. Например, у тысячелистника агрегатное соцветие - корзинки, собранные в щиток; у овсяницы, мятлика - колоски, собранные в метелку. Простые соцветия с удлиненной осью: · кисть - главная ось несет цветки с цветоножками, обычно одинаковой длины; · колос - главная ось несет цветки без цветоножек; · сережка - повислый колос, то есть колос с мягкой осью; после цветения соцветие обычно опадает; · початок - колос с сильно утолщенной осью, окружен одним или несколькими листьями, так называемым покрывалом, или крылом. Простые соцветия с укороченной осью: · зонтик - соцветие, у которого главная ось укорочена, и цветоножки, имеющие почти одинаковую длину, отходят от верхушки оси; · головка - зонтик, у которого цветки или без цветоножек, или очень короткие; · корзинка - верхушка главной оси разрастается в виде ложа, на котором расположены плотно сомкнутые цветки; верхушечные листья скучены и образуют обвертку. Цимоидные соцветия - это обширная группа соцветий. Среди цимоидных соцветий выделяют два основных типа: цимоиды и тирсы. Цимоиды, как правило, представляют собой упрощенные тирсы. Тирсы - это разветвленные соцветия, причем степень разветвления уменьшается от основания к верхушке. Главная ось тирса нарастает моноподиально, но парциальными соцветиями того или иного порядка являются цимоиды. Вопрос 11. Сущность и биологическое значение двойного оплодотворения. Явление апомиксиса. АПОМИКСИС (смешение), размножение растений, не сопровождающееся половым процессом. Термин «апомиксис» ввёл немецкий генетик Г. Винклер в 1908 году. В широком понимании явление апомиксис распространяют и на животных (многие перепончатокрылые, жуки, некоторые ящерицы, рыбы и др.), у которых встречаются формы нерегулярного полового размножения - партеногенез, гиногенез, андрогенез.
|
Последнее изменение этой страницы: 2017-05-11; Просмотров: 341; Нарушение авторского права страницы