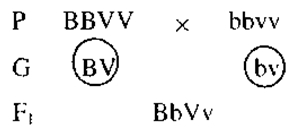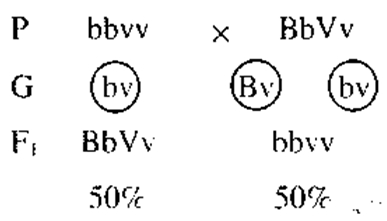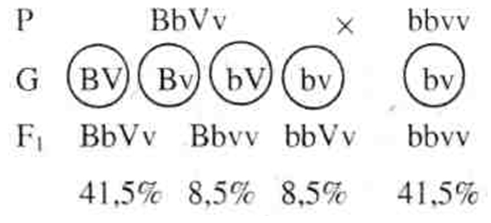|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Основные направления биотехнологииСтр 1 из 5Следующая ⇒
Задачи и методы селекции Селекция разрабатывает методы создания новых сортов растений, пород животных и штаммов микроорганизмов с необходимыми человеку признаками. Породы животных, сорта растений, штаммы микроорганизмов представляют собой совокупности особей, созданных человеком с помощью методов селекции, и характеризуются определенными наследственными особенностями, морфологическими и физиологическими хозяйственно ценными качествами. Поскольку свойства живых организмов обусловлены их нормой реакции на основе определенной генетической информации и подвержены модификационной и наследственной изменчивости, развитие селекции основано на закономерностях генетики. В селекционной работе используют следующие методы. Естественный отбор Стихийный (бессознательный). Проводился человеком для сохранения особей с наиболее ценными признаками, без стремления улучшить породу или сорт. Методический (сознательный). Характеризуется тем, что человек осознано и систематически стремится к выведению сорта или породы с желаемыми качествами. Подразделяется на: массовый — проводится по фенотипу. При этом отбирается группа особей, имеющих внешнее сходство. Поскольку генотипы отобранных особей по фенотипу неоднородны, отбор время от времени повторяют; индивидуальный — от каждой особи получают отдельное потомство и при последующих близкородственных скрещиваниях у животных и самоопылении у растений выводят чистые линии. Чистые линии — группы генетически однородных (гомозиготных) организмов представляют ценный исходный материал для селекции. Гибридизация Внутривидовая — основу составляет направленное скрещивание особей с интересующими селекционера свойствами и последующий отбор потомков с максимальным проявлением этих свойств. Близкородственная, или инбридинг, — проводится между братьями и сестрами или между родителями и потомством. В результате увеличивается доля гомозиготных организмов. В связи с переходом генов в гомозиготное состояние у потомков проявляются рецессивные мутации, что приводит к ослаблению потомства, появлению наследственных заболеваний. Межвидовая (отдаленная, неродственная) — проводится между особями разных видов и разных родов. Ее используют как селекционный метод, позволяющий объединить в гибриде ценные хозяйственные признаки родительских форм. В силу генетических, морфологических, физиологических и иных различий организмов разных видов отдаленная гибридизация, как правило, осуществляется с большим трудом и требует применения специальных методов преодоления нескрещиваемости. Межвидовые гибриды часто оказываются бесплодными вследствие нарушения процессов гаметогенеза, так как хромосомы, полученные от разных видов, различаются между собой и не конъюгируют, поэтому мейоз не происходит. Аутбридинг (скрещивание неродственных особей) — скрещивание особей разных линий. При данном скрещивании удается получить гетерозисные гибриды, превосходящие по своей мощности не только родительские линии, но и исходные формы, из которых эти линии были получены. Гетерозис заключается в повышенной мощности гибридов первого поколения по сравнению с родительскими формами. Основной причиной эффекта гетерозиса является отсутствие проявления вредных рецессивных аллелей в гетерозиготном состоянии. Полиплоидизация Увеличение числа наборов хромосом под действием на делящуюся клетку ядов, разрушающих веретено деления (например, колхицином). Индуцированный (искусственно вызванный) мутагенез В естественных условиях частота мутирования генов сравнительно невелика. Мутагенез повышает количество мутаций под воздействием на организм различных мутагенов: ультрафиолетовых лучей, ионизирующего излучения, некоторых химических веществ. Мутации в целом не носят направленный характер, селекционер отбирает и культивирует организмы с интересующими его признаками. Чаще всего используется в селекции микроорганизмов. В настоящее время активно развивается биотехнология — наука о способах получения необходимых человеку веществ с использованием живых организмов и биохимических процессов в производстве. ДНК-диагностика Это наиболее точный метод диагностики моногенных наследственных заболеваний. Преимущества метода: 1) позволяет определить причину заболевания на генетическом уровне; 2) выявляет минимальные нарушения структуры ДНК; 3) малоинвазивен; 4) не требует повторения. В основе метода лежит увеличение копий фрагментов ДНК различными способами.
Рис. 3. Наследственные заболевания, связанные с изменением числа хромосом: а — синдром Дауна (лишняя хромосома в 21-й паре); б — синдром Клайнфельтера (лишняя Х-хромосома у мужчины); в — синдром трисомии по Х-хромосомам (лишняя Х-хромосома у женщины); г — синдром Тернера-Шершевского (нехватка одной Х-хромосомы у женщины) Актуальные задачи медицинской генетики. Можно выделить несколько наиболее важных перспектив: > изучение причин возникновения мутаций, приводящих к наследственным заболеваниям и аномалиям развития у человека (разнородные химические и физические факторы); > поиск и разработка антимутагенных препаратов (предохраняющих от действия мутагенов); > профилактика и лечение наследственных болезней человека (разработка методов диагностики и поиски методов лечения); > развитие и расширение медико-генетического консультирования, которое должно помогать населению в прогнозировании здоровья будущих или родившихся детей; очень важна ранняя диагностика при беременности.
Сцепленное наследование. Генетика пола. В 1911 —1912 годах Т. Моргани сотрудники проверили проявление третьего закона Менделя на мухах-дрозофилах. Они учитывали две пары альтернативных признаков: серый (В) и черный (Ь) цвет тела и нормальные (V) и короткие (v) крылья. При скрещивании гомозиготных особей с серым цветом тела и нормальными крыльями с мухами с черным цветом тела и короткими крыльями получили единообразие гибридов первого поколения — мух с серым телом и нормальными крыльями. Подтвердился I закон Менделя.
Далее Морган решил провести анализирующее скрещивание гибридов первого поколения. Рецессивную гомозиготную самку он скрестил с дигетерозиготным самцом.
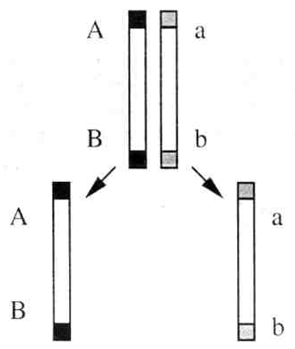
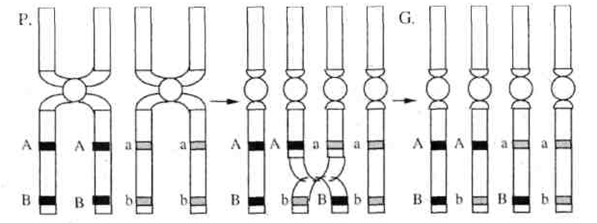
Морган ожидал получить, согласно третьему закону Менделя, мух четырех разных фенотипов в равном количестве (по 25%), а получил двух фенотипов (по 50% каждого). Морган пришел к выводу, что поскольку у организмов генов много, а хромосом относительно мало, то, следовательно, в каждой хромосоме содержится большое количество генов, и гены, локализованные в одной хромосоме, передаются вместе (сцепленно).Цитологические основы этого явления можно пояснить следующей схемой (рис. 1). Одна из пары гомологичных хромосом содержит два доминантных гена (BV), а другая — два рецессивных (bv). При мейозе хромосома с генами BV попадет в одну гамету, а хромосома с генами bv в другую.
Рис. 1. Схема расхождения гомологичных хромосом в мейозе при полном сцеплении. Таким образом, у дигетерозиготного организма образуются не четыре типа гамет (когда гены расположены в разных хромосомах), а только два, и, следовательно, потомки будут иметь два сочетания признаков (как у родителей). Гены, локализованные в одной хромосоме, обычно передаются вместе и составляют одну группу сцепления.Так как в гомологичных хромосомах локализованы аллельные гены, то группу сцепления составляют две гомологичные хромосомы, и, следовательно, количество групп сцеплениясоответствует количеству пар хромосом(или гаплоидному числу хромосом). Так, у мухи-дрозофилы всего 8 хромосом — 4 труппы сцепления, у человека 46 хромосом — 23 группы сцепления. Если гены, локализованные в одной хромосоме, передаются всегда вместе, то такое сцепление называется полным.Однако при дальнейшем анализе сцепления генов было обнаружено, что в некоторых случаях оно может нарушаться. Если дигетерозиготную самку мухи-дрозофилы скрестить с рецессивным самцом, результат будет следующий:
Морган предполагал получить опять мух четырех фенотипов по 25%, а получил потомков четырех фенотипов, но в другом соотношении: по 41, 5% особей с серым телом и нормальными крыльями и с черным телом и короткими крыльями и по 8, 5% мух с серым телом и короткими крыльями и с черным телом и нормальными крыльями. В этом случае сцепление генов неполное, т.е. гены, локализованные в одной хромосоме, не всегда передаются вместе.Это связано с явлением кроссинговера, которое заключается в обмене участками гомологичных хроматид в процессе их конъюгации в профазе мейоза I (рис. 2). Кроссинговер у гетерозиготных организмов приводит к перекомбинации генетического материала.
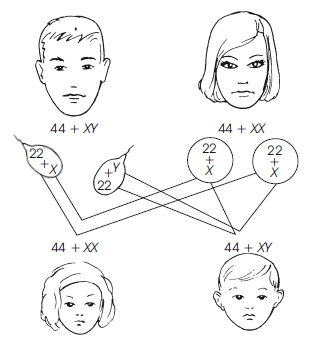
Рис. 2. Схема кроссинговера Каждая из образовавшихся хроматид попадает в отдельную гамету. Образуются 4 типа гамет, но в отличие от свободного комбинирования их процентное соотношение будет неравным, так как кроссинговер происходит не всегда. Частота кроссииговера зависит от расстояния между генами: чем больше расстояние, тем чаще может происходить кроссинговер. Расстояние между генами определяется в процентах кроссииговера — 1 морганидаравна 1 % кроссинговера. Итак, свободное комбинирование генов, согласно третьему закону Менделя, происходит в том случае, когда исследуемые гены расположены в разных хромосомах. Неполное сцепление наблюдается тогда, когда происходит перекомбинация генов (кроссинговер), расположенных в одной хромосоме. Если гены расположены в одной хромосоме и кроссинговер не происходит, сцепление будет полным. Кроссинговер имеет место у всех растений и животных, за исключением самца мухи-дрозофилы и самки тутового шелкопряда. Основные положения хромосомной теории наследственности: — гены расположены в хромосомах линейно в определенных локусах (участках); аллельные гены занимают одинаковые локусы в гомологичных хромосомах; —гены гомологичных хромосом образуют группу сцепления; число их равно гаплоидному набору хромосом; —между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер); —расстояние между генами пропорционально проценту кроссииговера и выражается в морганидах. Пол организма — это совокупность признаков и анатомических структур, обеспечивающих половой путь размножения и передачу наследственной информации. В определении пола будущей особи ведущую роль играет хромосомный аппарат зиготы — кариотип. Различают хромосомы, одинаковые для обоих полов — аутосомы, и половые хромосомы. В кариотипе человека содержится 44 аутосомы и 2 половых хромосомы — Х и Y. За развитие женского пола у человека отвечают две Х-хромосомы, т. е. женский пол гомогаметен. Развитие мужского пола определяется наличием Х- и Y-хромосом, т. е. мужской пол гетерогаметен. Сочетание половых хромосом в зиготе определяет пол будущего организма (рис. 3).
Рис. 3. Схема определения пола у человека. Половина сперматозоидов несетX-хромосому, а другая половина —Y-хромосому.Пол ребенка зависит от того, какой сперматозоид оплодотворитяйцеклетку У всех млекопитающих, человека и мухи-дрозофилы, гомогаметным является женский пол, а гетерогаметным — мужской. У птиц и бабочек, наоборот, гомогаметен мужской пол, а женский — гетерогаметен. Х-сцепленное наследование Так как Х-хромосома присутствует в кариотипе каждого человека, то и признаки, наследуемые сцеплено с Х-хромосомой, проявляются у представителей обоих полов. Женщины получают эти гены от обоих родителей и через свои гаметы передают их потомкам. Мужчины получают Х-хромосому от матери и передают ее своему потомству женского пола. Различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. У человека Х-сцепленный доминантный признак передается матерью всему потомству. Мужчина передает свой Х-сцепленный доминантный признак лишь своим дочерям. Х-сцепленный рецессивный признак у женщин проявляется лишь при получении ими соответствующего аллеля от обоих родителей. У мужчин он развивается при получении рецессивного аллеля от матери. Женщины передают рецессивный аллель потомкам обоих полов, а мужчины — только дочерям. При Х-сцепленном наследовании возможен промежуточный характер проявления признака у гетерозигот. Y-сцепленные гены присутствуют в генотипе только мужчин и передаются из поколения в поколение от отца к сыну.
Неаллельные гены. Наследственность. Неаллельные гены Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы взаимодействия неаллельных генов: 1) комплементарность; 2) эпистаз; 3) полимерия. Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9: 6: 1, 9: 3: 4, 9: 7, иногда 9: 3: 3: 1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлиненную. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдет расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлиненные. Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (B, b) подавляется доминантным эпистатичным геном (I > B, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12: 3: 1, 13: 3, 7: 6: 3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > B, b). Расщепление по фенотипу может идти в соотношении 9: 3: 4, 9: 7, 13: 3. Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 по фенотипу происходит в соотношении 1: 4: 6: 4: 1. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15: 1.
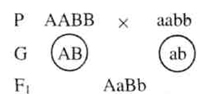
Взаимодействия аллельных генов. Наследование групп крови системы АВО Взаимодействия аллельных генов При взаимодействии аллельных генов возможны разные варианты проявления признака. Если аллели находятся в гомозиготном состоянии, то развивается соответствующий аллелю вариант признака. В случае гетерозиготности развитие признака будет зависеть от конкретного вида взаимодействия аллельных генов. Полное доминирование Это такой вид взаимодействия аллельных генов, при котором проявление одного из аллелей (А) не зависит от наличия в генотипе особи другого аллеля (А1) и гетерозиготы АА1 фенотипиче-ски не отличаются от гомозигот по данному аллелю (АА). В гетерозиготном генотипе АА1 аллель А является доминантным. Присутствие аллеля А1 никак фенотипически не проявляется, поэтому он выступает как рецессивный. Неполное доминирование Отмечается в случаях, когда фенотип гетерозигот СС1 отличается от фенотипа гомозигот СС и С1С1 промежуточной степенью проявления признака, т. е. аллель, отвечающий за формирование нормального признака, находясь в двойной дозе у гомозиготы СС, проявляется сильнее, чем в одинарной дозе у гетерозиготы СС1. Возможные при этом генотипы различаются экспрессивностью, т. е. степенью выраженности признака. Кодоминирование Это такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате формируется промежуточный вариант признака, новый по сравнению с вариантами, формируемыми каждым аллелем по отдельности. Межаллельная комплементация Это редкий вид взаимодействия аллельных генов, при котором у организма, гетерозиготного по двум мутантным аллелям гена М (М1М11), возможно формирование нормального признака М. Например, ген М отвечает за синтез белка, имеющего четвертичную структуру и состоящего из нескольких одинаковых полипептидных цепей. Мутантный аллель М1 вызывает синтез измененного пептида М1, а мутантный аллель М11 определяет синтез другой, но тоже ненормальной полипептидной цепи. Взаимодействие таких измененных пептидов и компенсация измененных участков при формировании четвертичной структуры в редких случаях может привести к появлению белка с нормальными свойствами. Наследование групп крови системы АВО Наследование групп крови системы АВО у человека имеет некоторые особенности. Формирование I, II и III групп крови происходит по такому типу взаимодействия аллельных генов, как доминирование. Генотипы, содержащие аллель IA в гомозиготном состоянии, либо в сочетании с аллелем IO, определяют формирование у человека второй (А) группы крови. Тот же принцип лежит в основе формирования третьей (В) группы крови, т. е. аллели IA и IB выступают как доминантные по отношению к аллелю IO, в гомозиготном состоянии формирующему IOIO первую (О) группу крови. Формирование четвертой (АВ) группы крови идет по пути кодоминирования. Аллели IA и IB, по отдельности формирующие соответственно вторую и третью группу крови, в гетерозиготном состоянии определяют IAIB (четвертую) группу крови. Ди- и полигибридное скрещивание. III закон Г. Менделя Изучив наследование одной пары аллелей, Мендель проследил наследование двух признаков одновременно. С этой целью он использовал гомозиготные растения гороха, отличающиеся по двум парам альтернативных признаков: семена желтые гладкие (А, В — доминантные признаки) и зеленые морщинистые (a, b — рецессивные признаки).
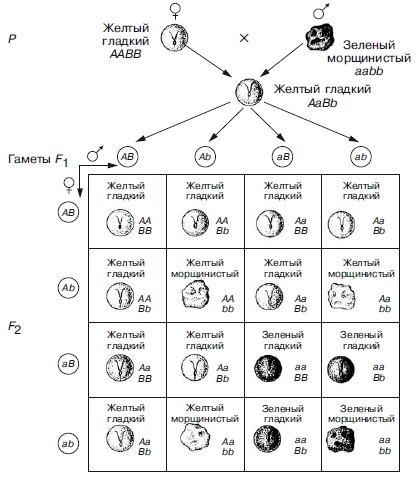
В результате такого скрещивания в первом поколении он получил растения, у которых все семена были желтые гладкие. Этот результат подтверждает, что закон единообразия гибридов первого поколения проявляется не только при моногибридном скрещивании, но и при дигибридном. Полученные гибриды первого поколения (АаВЬ) будут давать четыре типа гамет в равном соотношении, так как в процессе мейоза из каждой пары генов в гамету попадает один ген, свободно комбинируясь с генами другой пары. При оплодотворении каждая из четырех типов гамет одного организма случайно встречается с одной из гамет другого. Следовательно, возможно 16 вариантов их сочетаний. Для удобства записи пользуются решеткой Пеннета, в которой по горизонтали записывают женские гаметы, а по вертикали — мужские:
Краткая запись генотипа (А-В-) применяется для обозначения фенотипа особи, так как независимо от второй аллели (А или а) фенотип особи будет доминантный (желтый). Легко подсчитать, что по фенотипу потомство делится на 4 группы: 9 частей растений с желтыми гладкими семенами (А-В-), 3 части — с желтыми морщинистыми (A-bb), 3 части с зелеными гладкими (ааВ-) и 1 часть — с зелеными морщинистыми (aabb). Если учесть расщепление по одной паре признаков (желтый и зеленый цвет, гладкая и морщинистая поверхность), то получится: 9 + 3 особи с желтыми (гладкими) и 3 + 1 особи с зелеными (морщинистыми) семенами. Их соотношение равно 12: 4, или 3: 1. Следовательно, при дигибридном скрещивании каждая пара признаков в потомстве даст расщепление независимо от другой пары, как и при моногибридном скрещивании. При этом происходит случайное комбинирование генов, приводящее к новым сочетаниям признаков, которых не было у родительских форм. В нашем примере исходные растения гороха имели желтые гладкие и зеленые морщинистые семена, а во втором поколении, кроме таких сочетаний признаков, получены растения с желтыми морщинистыми и зелеными гладкими семенами.
Рис. 1. Схема наследования окраски и формы семян у гороха: А — желтая, а — зеленая; В — гладкая, b — морщинистая Отсюда следует третийзакон Менделя— закон независимого наследования двух разных признаков: при скрещивании гомозиготных особей, анализируемых по двум или нескольким парам альтернативных признаков, во втором поколении наблюдается независимое комбинирование генов разных аллельных пар и соответствующих им признаков. Закон базируется на независимом расхождении в мейозе гомологичных хромосом разных пар. Опыты Менделя легли в основу новой науки — генетики. Генетика — это наука, изучающая наследственность и изменчивость. Успеху исследований Менделя способствовали следующие условия: 1. Удачный выбор объекта исследования — гороха. Когда Менделю предложили повторить свои наблюдения на ястребинке, этом вездесущем сорняке, он не смог этого сделать. 2. Проведение анализа наследования отдельных пар признаков в потомстве скрещиваемых растений, отличающихся по одной, двум или трем парам альтернативных признаков. Велся учет отдельно по каждой паре этих признаков после каждого скрещивания. 3. Мендель не только зафиксировал полученные результаты, но и провел их математический анализ. Мендель сформулировал также закон чистоты гамет, согласно которому гамета чиста от второго аллельного гена (альтернатив ного признака), т. е. ген дискретен и не смешивается с другими генами. При моногибридном скрещивании в случае полного доминирования у гетерозиготных гибридов первого поколения проявляется только доминантный аллель, однако рецессивный аллель не теряется и не смешивается с доминантным. Среди гибридов второго поколения и рецессивный, и доминантный аллель может проявиться в своем — чистом — виде, т. е. в гомозиготном состоянии. В итоге гаметы, образуемые такой гетерозиготой, являются чистыми, т. е. гамета А не содержит ничего от аллели а, гамета а — чиста от А. На клеточном уровне основой дискретности аллелей является их локализация в разных хромосомах каждой гомологичной пары, а дискретности генов — их расположение в разных локусах хромосом.
Закономерности наследования, установленные Г. Менделем Моногибридное скрещивание. Некоторые закономерности наследования были впервые установлены Г. Менделем. Он достиг успеха в своих экспериментах благодаря использованию гибридологического метода —скрещивания организмов, различающихся по каким-либо признакам, и анализа всех последующих поколений с целью установления закономерностей наследования этих признаков. Гибридологический метод и до настоящего времени остается одним из основных в генетических исследованиях. Г. Мендель усовершенствовал данный метод, и в отличие от своих предшественников, анализировал наследование ограниченного количества признаков (одного, двух, трех). При этом он выбирал признак с альтернативным(контрастирующим) проявлением его у скрещиваемых организмов. Так, он скрещивал сорта гороха с окрашенными и белыми цветками, гладкими и морщинистыми семенами и т. п. Кроме того, Мендель проверял перед скрещиванием, насколько устойчиво наследуются выбранные им признаки в ряду поколений при самоопылении. В процессе эксперимента им проводился также точный количественный учет всех гибридных растений во всех поколениях. Основные понятия современной генетики. Наследственностью называется свойство организмов повторять в ряду поколений комплекс признаков (особенности внешнего строения, физиологии, химического состава, характера обмена веществ, индивидуального развития и т. д.). Изменчивость — явление, противоположное наследственности. Она заключается в изменении комбинаций признаков или появлении совершенно новых признаков у особей данного вида. Благодаря наследственности обеспечивается сохранение видов на протяжении значительных промежутков (до сотен миллионов лет) времени. Однако условия окружающей среды меняются (иногда существенно) с течением времени, и в таких случаях изменчивость, приводящая к разнообразию особей внутри вида, обеспечивает его выживание. Какие-то из особей оказываются более приспособленными к новым условиям, это и позволяет им выжить. Кроме того, изменчивость позволяет видам расширять границы своего местообитания, осваивать новые территории. Сочетание двух указанных свойств тесно связано с процессом эволюции. Новые признаки организмов появляются в результате изменчивости, а благодаря наследственности они сохраняются в последующих поколениях. Накапливание множества новых признаков приводит к возникновению других видов Виды изменчивости Различают наследственную и ненаследственную изменчивость. Наследственная (генотипическая) изменчивость связана с изменением самого генетического материала. Ненаследственная (фенотипическая, модификационная) изменчивость — это способность организмов изменять свой фенотип под влиянием различных факторов. Причиной модификационной изменчивости являются изменения внешней среды обитания организма или его внутренней среды. Норма реакции Это границы фенотипической изменчивости признака, возникающей под действием факторов внешней среды. Норма реакции определяется генами организма, поэтому норма реакции по одному и тому же признаку у разных индивидов различна. Размах нормы реакции различных признаков также варьирует. Те организмы, у которых норма реакции шире по данному признаку, обладают более высокими адаптивными возможностями в определенных условиях среды, т. е. модификационная изменчивость в большинстве случаев носит адаптивный характер, и большинство изменений, возникших в организме при воздействии определенных факторов внешней среды, являются полезными. Однако фенотипические изменения иногда утрачивают приспособительный характер. Если фенотипическая изменчивость клинически сходна с наследственным заболеванием, то такие изменения называются фенокопией. Комбинативная изменчивость Связана с новым сочетанием неизменных генов родителей в генотипах потомства. Факторы комбинативной изменчивости. 1.Независимое и случайное расхождение гомологичных хромосом в анафазе I мейоза. 2.Кроссинговер. 3.Случайное сочетание гамет при оплодотворении. 4.Случайный подбор родительских организмов. Мутации Это редкие, случайно возникшие стойкие изменения генотипа, затрагивающие весь геном, целые хромосомы, части хромосом или отдельные гены. Они возникают под действием мутагенных факторов физического, химического или биологического происхождения. Мутации бывают: 1) спонтанные и индуцированные; 2) вредные, полезные и нейтральные; 3) соматические и генеративные; 4) генные, хромосомные и геномные. Спонтанные мутации — это мутации, возникшие ненаправленно, под действием неизвестного мутагена. Индуцированные мутации — это мутации, вызванные искусственно действием известного мутагена. Хромосомные мутации — это изменения структуры хромосом в процессе клеточного деления. Различают следующие виды хромосомных мутаций. 1.Дупликация — удвоение участка хромосомы за счет неравного кроссинговера. 2.Делеция — потеря участка хромосомы. 3.Инверсия — поворот участка хромосомы на 180°. 4.Транслокация — перемещение участка хромосомы на другую хромосому. Геномные мутации — это изменение числа хромосом. Виды геномных мутаций. 1.Полиплоидия — изменение числа гаплоидных наборов хромосом в кариотипе. Под кариотипом понимают число, форму и количество хромосом, характерные для данного вида. Различают нуллисомию (отсутствие двух гомологичных хромосом), моносомию (отсутствие одной из гомологичных хромосом) и полисомию (наличие двух и более лишних хромосом). 2.Гетероплоидия — изменение числа отдельных хромосом в кариотипе. Генные мутации встречаются наиболее часто. Причины генных мутаций: 1) выпадение нуклеотида; 2) вставка лишнего нуклеотида (эта и предыдущая причины приводят к сдвигу рамки считывания); 3) замена одного нуклеотида на другой. Передача наследственных признаков в ряду поколений особей осуществляется в процессе размножения. При половом — через половые клетки, при бесполом наследственные признаки передаются с соматическими клетками. Единицами наследственности (ее материальными носителями) являются гены. В функциональном отношении конкретный ген отвечает за развитие какого-то признака. Это не противоречит тому определению, которое мы давали гену выше. С химической точки зрения ген - участок молекулы ДНК. Он содержит генетическую информацию о структуре синтезируемого белка (т. е. последовательности аминокислот в белковой молекуле). Совокупность всех генов в организме определяет совокупность конкретных белков, синтезируемых в нем, что в конечном счете приводит к формированию специфических признаков. У прокариотной клетки гены входят в состав единственной молекулы ДНК, а у эукариотной - в молекулы ДНК, заключенные в хромосомах. При этом в паре гомологичных хромосом в одних и тех же участках располагаются гены, отвечающие за развитие какого-то признака (например, окраска цветка, форма семян, цвет глаз у человека). Они получили название аллельных генов. В одну пару аллельных генов могут входить либо одинаковые (по составу нуклеотидов и определяемому ими признаку), либо отличающиеся гены. Понятие «признак» связано с каким-то отдельным качеством организма (морфологическим, физиологическим, биохимическим), по которому мы можем отличить его от другого организма. Например: глаза голубые или карие, цветки окрашенные или неокрашенные, рост высокий или низкий, группа крови I(0) или II(A) и т. д. Совокупность всех генов у организма называется генотипом, а совокупность всех признаков — фенотипом. Фенотип формируется на базе генотипа в определенных условиях внешней среды в ходе индивидуального развития организмов.
Задачи и методы селекции Селекция разрабатывает методы создания новых сортов растений, пород животных и штаммов микроорганизмов с необходимыми человеку признаками. Породы животных, сорта растений, штаммы микроорганизмов представляют собой совокупности особей, созданных человеком с помощью методов селекции, и характеризуются определенными наследственными особенностями, морфологическими и физиологическими хозяйственно ценными качествами. Поскольку свойства живых организмов обусловлены их нормой реакции на основе определенной генетической информации и подвержены модификационной и наследственной изменчивости, развитие селекции основано на закономерностях генетики. В селекционной работе используют следующие методы. Естественный отбор Стихийный (бессознательный). Проводился человеком для сохранения особей с наиболее ценными признаками, без стремления улучшить породу или сорт. Методический (сознательный). Характеризуется тем, что человек осознано и систематически стремится к выведению сорта или породы с желаемыми качествами. Подразделяется на: массовый — проводится по фенотипу. При этом отбирается группа особей, имеющих внешнее сходство. Поскольку генотипы отобранных особей по фенотипу неоднородны, отбор время от времени повторяют; индивидуальный — от каждой особи получают отдельное потомство и при последующих близкородственных скрещиваниях у животных и самоопылении у растений выводят чистые линии. Чистые линии — группы генетически однородных (гомозиготных) организмов представляют ценный исходный материал для селекции. Гибридизация Внутривидовая — основу составляет направленное скрещивание особей с интересующими селекционера свойствами и последующий отбор потомков с максимальным проявлением этих свойств. Близкородственная, или инбридинг, — проводится между братьями и сестрами или между родителями и потомством. В результате увеличивается доля гомозиготных организмов. В связи с переходом генов в гомозиготное состояние у потомков проявляются рецессивные мутации, что приводит к ослаблению потомства, появлению наследственных заболеваний. Межвидовая (отдаленная, неродственная) — проводится между особями разных видов и разных родов. Ее используют как селекционный метод, позволяющий объединить в гибриде ценные хозяйственные признаки родительских форм. В силу генетических, морфологических, физиологических и иных различий организмов разных видов отдаленная гибридизация, как правило, осуществляется с большим трудом и требует применения специальных методов преодоления нескрещиваемости. Межвидовые гибриды часто оказываются бесплодными вследствие нарушения процессов гаметогенеза, так как хромосомы, полученные от разных видов, различаются между собой и не конъюгируют, поэтому мейоз не происходит. Аутбридинг (скрещивание неродственных особей) — скрещивание особей разных линий. При данном скрещивании удается получить гетерозисные гибриды, превосходящие по своей мощности не только родительские линии, но и исходные формы, из которых эти линии были получены. Гетерозис заключается в повышенной мощности гибридов первого поколения по сравнению с родительскими формами. Основной причиной эффекта гетерозиса является отсутствие проявления вредных рецессивных аллелей в гетерозиготном состоянии. Полиплоидизация Увеличение числа наборов хромосом под действием на делящуюся клетку ядов, разрушающих веретено деления (например, колхицином). Индуцированный (искусственно вызванный) мутагенез |
Последнее изменение этой страницы: 2017-05-04; Просмотров: 47; Нарушение авторского права страницы