|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Признаки, сцепленные с полом
Это признаки, которые кодируются генами, находящимися на половых хромосомах. У человека признаки, кодируемые генами Х-хромосомы, могут проявляться у представителей обоих полов, а кодируемые генами Y-хромосомы — только у мужчин. Следует иметь в виду, что в мужском генотипе только одна Х-хромосома, которая почти не содержит участков, гомологичных с Y-хромосомой, поэтому все локализованные в Х-хромосоме гены, в том числе и рецессивные, проявляются в фенотипе в первом же поколении. В половых хромосомах содержатся гены, регулирующие проявление не только половых признаков. Х-хромосома имеет гены, отвечающие за свертываемость крови, цветовое восприятие, синтез ряда ферментов. В Y-хромосоме содержится ряд генов, контролирующих признаки, наследуемые по мужской линии (голандрические признаки): волосистость ушной раковины, наличие кожной перепонки между пальцами и др. Известно очень мало генов, общих для Х- и Y-хромосом. Различают Х-сцепленное и Y-сцепленное (голандрическое) наследование. Х-сцепленное наследование Так как Х-хромосома присутствует в кариотипе каждого человека, то и признаки, наследуемые сцеплено с Х-хромосомой, проявляются у представителей обоих полов. Женщины получают эти гены от обоих родителей и через свои гаметы передают их потомкам. Мужчины получают Х-хромосому от матери и передают ее своему потомству женского пола. Различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. У человека Х-сцепленный доминантный признак передается матерью всему потомству. Мужчина передает свой Х-сцепленный доминантный признак лишь своим дочерям. Х-сцепленный рецессивный признак у женщин проявляется лишь при получении ими соответствующего аллеля от обоих родителей. У мужчин он развивается при получении рецессивного аллеля от матери. Женщины передают рецессивный аллель потомкам обоих полов, а мужчины — только дочерям. При Х-сцепленном наследовании возможен промежуточный характер проявления признака у гетерозигот. Y-сцепленные гены присутствуют в генотипе только мужчин и передаются из поколения в поколение от отца к сыну.
Неаллельные гены. Наследственность. Неаллельные гены Неаллельные гены — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой. При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы взаимодействия неаллельных генов: 1) комплементарность; 2) эпистаз; 3) полимерия. Комплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9: 6: 1, 9: 3: 4, 9: 7, иногда 9: 3: 3: 1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлиненную. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдет расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлиненные. Эпистаз — взаимодействие неаллельных генов, при котором один из них подавляется другим. Подавляющий ген называется эпистатичным, подавляемый — гипостатичным. Если эпистатичный ген не имеет собственного фенотипического проявления, то он называется ингибитором и обозначается буквой I. Эпистатическое взаимодействие неаллельных генов может быть доминантным и рецессивным. При доминантном эпистазе проявление гипостатичного гена (B, b) подавляется доминантным эпистатичным геном (I > B, b). Расщепление по фенотипу при доминантном эпистазе может происходить в соотношении 12: 3: 1, 13: 3, 7: 6: 3. Рецессивный эпистаз — это подавление рецессивным аллелем эпистатичного гена аллелей гипостатичного гена (i > B, b). Расщепление по фенотипу может идти в соотношении 9: 3: 4, 9: 7, 13: 3. Полимерия — взаимодействие неаллельных множественных генов, однозначно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммирующего действия генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление F2 по фенотипу происходит в соотношении 1: 4: 6: 4: 1. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление по фенотипу происходит в соотношении 15: 1.
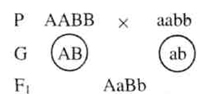
Взаимодействия аллельных генов. Наследование групп крови системы АВО Взаимодействия аллельных генов При взаимодействии аллельных генов возможны разные варианты проявления признака. Если аллели находятся в гомозиготном состоянии, то развивается соответствующий аллелю вариант признака. В случае гетерозиготности развитие признака будет зависеть от конкретного вида взаимодействия аллельных генов. Полное доминирование Это такой вид взаимодействия аллельных генов, при котором проявление одного из аллелей (А) не зависит от наличия в генотипе особи другого аллеля (А1) и гетерозиготы АА1 фенотипиче-ски не отличаются от гомозигот по данному аллелю (АА). В гетерозиготном генотипе АА1 аллель А является доминантным. Присутствие аллеля А1 никак фенотипически не проявляется, поэтому он выступает как рецессивный. Неполное доминирование Отмечается в случаях, когда фенотип гетерозигот СС1 отличается от фенотипа гомозигот СС и С1С1 промежуточной степенью проявления признака, т. е. аллель, отвечающий за формирование нормального признака, находясь в двойной дозе у гомозиготы СС, проявляется сильнее, чем в одинарной дозе у гетерозиготы СС1. Возможные при этом генотипы различаются экспрессивностью, т. е. степенью выраженности признака. Кодоминирование Это такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате формируется промежуточный вариант признака, новый по сравнению с вариантами, формируемыми каждым аллелем по отдельности. Межаллельная комплементация Это редкий вид взаимодействия аллельных генов, при котором у организма, гетерозиготного по двум мутантным аллелям гена М (М1М11), возможно формирование нормального признака М. Например, ген М отвечает за синтез белка, имеющего четвертичную структуру и состоящего из нескольких одинаковых полипептидных цепей. Мутантный аллель М1 вызывает синтез измененного пептида М1, а мутантный аллель М11 определяет синтез другой, но тоже ненормальной полипептидной цепи. Взаимодействие таких измененных пептидов и компенсация измененных участков при формировании четвертичной структуры в редких случаях может привести к появлению белка с нормальными свойствами. Наследование групп крови системы АВО Наследование групп крови системы АВО у человека имеет некоторые особенности. Формирование I, II и III групп крови происходит по такому типу взаимодействия аллельных генов, как доминирование. Генотипы, содержащие аллель IA в гомозиготном состоянии, либо в сочетании с аллелем IO, определяют формирование у человека второй (А) группы крови. Тот же принцип лежит в основе формирования третьей (В) группы крови, т. е. аллели IA и IB выступают как доминантные по отношению к аллелю IO, в гомозиготном состоянии формирующему IOIO первую (О) группу крови. Формирование четвертой (АВ) группы крови идет по пути кодоминирования. Аллели IA и IB, по отдельности формирующие соответственно вторую и третью группу крови, в гетерозиготном состоянии определяют IAIB (четвертую) группу крови. Ди- и полигибридное скрещивание. III закон Г. Менделя Изучив наследование одной пары аллелей, Мендель проследил наследование двух признаков одновременно. С этой целью он использовал гомозиготные растения гороха, отличающиеся по двум парам альтернативных признаков: семена желтые гладкие (А, В — доминантные признаки) и зеленые морщинистые (a, b — рецессивные признаки).
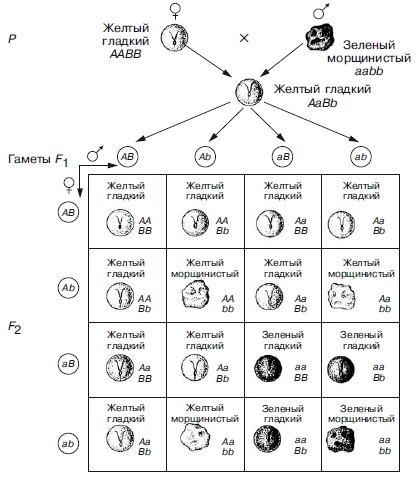
В результате такого скрещивания в первом поколении он получил растения, у которых все семена были желтые гладкие. Этот результат подтверждает, что закон единообразия гибридов первого поколения проявляется не только при моногибридном скрещивании, но и при дигибридном. Полученные гибриды первого поколения (АаВЬ) будут давать четыре типа гамет в равном соотношении, так как в процессе мейоза из каждой пары генов в гамету попадает один ген, свободно комбинируясь с генами другой пары. При оплодотворении каждая из четырех типов гамет одного организма случайно встречается с одной из гамет другого. Следовательно, возможно 16 вариантов их сочетаний. Для удобства записи пользуются решеткой Пеннета, в которой по горизонтали записывают женские гаметы, а по вертикали — мужские:
Краткая запись генотипа (А-В-) применяется для обозначения фенотипа особи, так как независимо от второй аллели (А или а) фенотип особи будет доминантный (желтый). Легко подсчитать, что по фенотипу потомство делится на 4 группы: 9 частей растений с желтыми гладкими семенами (А-В-), 3 части — с желтыми морщинистыми (A-bb), 3 части с зелеными гладкими (ааВ-) и 1 часть — с зелеными морщинистыми (aabb). Если учесть расщепление по одной паре признаков (желтый и зеленый цвет, гладкая и морщинистая поверхность), то получится: 9 + 3 особи с желтыми (гладкими) и 3 + 1 особи с зелеными (морщинистыми) семенами. Их соотношение равно 12: 4, или 3: 1. Следовательно, при дигибридном скрещивании каждая пара признаков в потомстве даст расщепление независимо от другой пары, как и при моногибридном скрещивании. При этом происходит случайное комбинирование генов, приводящее к новым сочетаниям признаков, которых не было у родительских форм. В нашем примере исходные растения гороха имели желтые гладкие и зеленые морщинистые семена, а во втором поколении, кроме таких сочетаний признаков, получены растения с желтыми морщинистыми и зелеными гладкими семенами.
Рис. 1. Схема наследования окраски и формы семян у гороха: А — желтая, а — зеленая; В — гладкая, b — морщинистая Отсюда следует третийзакон Менделя— закон независимого наследования двух разных признаков: при скрещивании гомозиготных особей, анализируемых по двум или нескольким парам альтернативных признаков, во втором поколении наблюдается независимое комбинирование генов разных аллельных пар и соответствующих им признаков. Закон базируется на независимом расхождении в мейозе гомологичных хромосом разных пар. Опыты Менделя легли в основу новой науки — генетики. Генетика — это наука, изучающая наследственность и изменчивость. Успеху исследований Менделя способствовали следующие условия: 1. Удачный выбор объекта исследования — гороха. Когда Менделю предложили повторить свои наблюдения на ястребинке, этом вездесущем сорняке, он не смог этого сделать. 2. Проведение анализа наследования отдельных пар признаков в потомстве скрещиваемых растений, отличающихся по одной, двум или трем парам альтернативных признаков. Велся учет отдельно по каждой паре этих признаков после каждого скрещивания. 3. Мендель не только зафиксировал полученные результаты, но и провел их математический анализ. Мендель сформулировал также закон чистоты гамет, согласно которому гамета чиста от второго аллельного гена (альтернатив ного признака), т. е. ген дискретен и не смешивается с другими генами. При моногибридном скрещивании в случае полного доминирования у гетерозиготных гибридов первого поколения проявляется только доминантный аллель, однако рецессивный аллель не теряется и не смешивается с доминантным. Среди гибридов второго поколения и рецессивный, и доминантный аллель может проявиться в своем — чистом — виде, т. е. в гомозиготном состоянии. В итоге гаметы, образуемые такой гетерозиготой, являются чистыми, т. е. гамета А не содержит ничего от аллели а, гамета а — чиста от А. На клеточном уровне основой дискретности аллелей является их локализация в разных хромосомах каждой гомологичной пары, а дискретности генов — их расположение в разных локусах хромосом.
Закономерности наследования, установленные Г. Менделем Моногибридное скрещивание. Некоторые закономерности наследования были впервые установлены Г. Менделем. Он достиг успеха в своих экспериментах благодаря использованию гибридологического метода —скрещивания организмов, различающихся по каким-либо признакам, и анализа всех последующих поколений с целью установления закономерностей наследования этих признаков. Гибридологический метод и до настоящего времени остается одним из основных в генетических исследованиях. Г. Мендель усовершенствовал данный метод, и в отличие от своих предшественников, анализировал наследование ограниченного количества признаков (одного, двух, трех). При этом он выбирал признак с альтернативным(контрастирующим) проявлением его у скрещиваемых организмов. Так, он скрещивал сорта гороха с окрашенными и белыми цветками, гладкими и морщинистыми семенами и т. п. Кроме того, Мендель проверял перед скрещиванием, насколько устойчиво наследуются выбранные им признаки в ряду поколений при самоопылении. В процессе эксперимента им проводился также точный количественный учет всех гибридных растений во всех поколениях. |
Последнее изменение этой страницы: 2017-05-04; Просмотров: 44; Нарушение авторского права страницы