|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Анаэробный распад углеводов в тканях. Последовательность реакций. Регуляция. Гликолитическая оксиредукция. Молочнокислое и другие виды брожения. Значение анаэробного распада глюкозы в организме.
Анаэробный распад глюкозы происходит при недостаточном содержании кислорода, в клетках мышечной ткани животного организма. Данный путь распада называется дихотомическим, т.к. в процессе происходит образование двух молекул триоз, содержащих по 3 С-атома из одной молекулы гексозы (6 С-атомов). Конечный продукт анаэробного превращения глюкозы – молочная кислота. Гликолиз протекает в гиалоплазме (цитозоле) клетки. Гликолиз условно можно разбить на два этапа. В первом этапе происходит затрата энергии, второй этап, наоборот, характеризуется накоплением энергии в форме молекул АТФ.Следует отметить, что в организме любой метаболический путь начинается с активации исходного соединения.Первой ферментативной реакцией гликолиза является фосфорилирование глюкозы, т.е. перенос остатка фосфорной кислоты на глюкозу за счет энергии АТФ с образованием глюкозо-6-фосфата. Реакция катализируется ферментом гексокиназой. Киназами называются ферменты, катализирующие перенос остатка фосфорной кислоты от АТФ к акцептору.
Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии и может считаться практически необратимым процессом. Наиболее важным свойством гексокиназы являетвся ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором. Второй реакцией гликолиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфат-изомеразы во фруктозо-6-фосфат (изомеризация альдозы в кетозу):
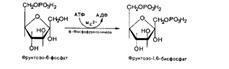
Эта реакция протекает легко в обоих направлениях. Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом. Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза. Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бифосфат расщепляется на две фосфотриозы:
Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры peакция сдвигается в сторону большего образования триозофосфатов (дигидроксиацетонфосфата и глицеральдегид-3-фосфата (фосфоглицериновый альдегид)). Пятая реакция - это реакция изомеризации триозофосфатов. Kaтализируется ферментом триозофосфатизомеразой:
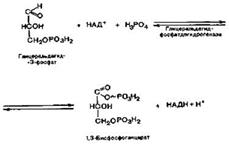
Дальнейшим превращениям будет подвергаться только глицеральдегид-3-фосфат. В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается окислению с образованием 1,3- бифосфоглицериновой кислоты и восстановленной формы НАДН. С данной реакции начинается второй этап гликолиза.
1,3-Бисфосфоглицерат представляет собой высокоэнергетическое соединение (макроэргическая связь условно обозначена знаком «тильда» ~). Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфоглицериноой кислоты (3-фосфоглицерат):
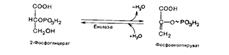
Это первая реакция гликолиза, в которой происходит образование АТФ (пример субстратного фосфорилирования). Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат):
Девятая реакция катализируется ферментом енолазой, при этом фосфоглицериновая кислота в результате отщепления молекулы переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстатное фосфолирование). Катализируется ферментом пируваткиназой:
В результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции:
Конечным акцептором электронов в ходе гликолиза является ПВК, которая восстанавливается в молочную кислоту (лактат). Поэтому для нормального протекания этого метаболического пути требуется только глюкоза, которая может быть запасена клеткой в виде резервных полисахаридов в избыточном количестве. Образование лактата является завершающей стадией анаэробного гликолиза. Энергетический баланс – 2 молекулы АТФ (4 молекулы образуется, 2 - потребляется). Впервые биологическую природу брожения открыл в 60-х годах 19 в. гениальный французский ученый Луи Пастер. Пастеру удалось на примере молочнокислого, спиртового и маслянокислого брожения доказать, что эти процессы вызываются жизнедеятельностью микроорганизмов. Спиртовое брожение углеводов вызывают дрожжи (Saccharomyces cerevisiae), некоторые виды бактерий (Sarcina ventriculi) и отдельные представители мукоровых грибов рода Mucor. При спиртовом брожении молекула гексозы распадается на этанол и углекислый газ. В ходе брожения образуется много промежуточных продуктов — гексозомонофосфат, фруктозодифосфат, фосфотриозы, фосфоглицериновая кислота, фосфопировиноградная кислота, пировиноградная кислота, уксусный альдегид и, наконец, этиловый спирт. При содержании в сбраживаемом растворе более чем 30% сахара часть его остается неиспользованной, так как при этих условиях образуется до 15% спирта, а при такой концентрации спирт подавляет жизнедеятельность дрожжей. Поэтому натуральные вина содержат не более 15% спирта. Главное преимущество чистых культур дрожжей заключается в том, что брожение виноградного сока протекает и заканчивается быстро, а отсутствие посторонней микрофлоры позволяет получать вина хорошего вкуса и аромата (с хорошим «букетом»). По окончании брожения молодое вино стабилизируют и дают ему созреть. Эти процессы занимают несколько месяцев, а при изготовлении высококачественных красных вин — даже несколько лет. В течение первого года во многих красных винах происходит второе, спонтанное брожение — яблочно-молочнокислое, которое вызывается рядом молочнокислых бактерий (Prdiococcus, Leuconostoc). В результате этого яблочная кислота винограда превращается в молочную кислоту и СО2, т. е. дикарбоновая кислота превращается в монокарбоновую, и кислотность вина уменьшается, оно становится высококачественным. Уксуснокислое брожение — биологический окислительный процесс, при котором с помощью уксуснокислых бактерий спирт окисляется в уксусную кислоту. Если какую-либо жидкость, содержащую небольшое количество спирта (вино, пиво), оставить открытой, то в ней постепенно появляется уксусная кислота и кожистая пленка (уксусная матка) на поверхности. Уксуснокислые бактерии объединены в род Acetobacter, содержащий ряд видов и подвидов. Этиловый спирт под влиянием уксуснокислых бактерий подвергается окислению, в результате которого вначале образуется уксусный альдегид, а затем — уксусная кислота. При использовании специальных рас уксуснокислых бактерий максимальный выход уксуса достигает 14,5%. Уксуснокислые бактерии превращают ряд многоатомных спиртов в сахар. Одна из таких реакций используется для получения сорбозы из сорбитола. Сорбоза — промежуточный продукт синтеза аскорбиновой кислоты. Она применяется в качестве суспендирующего агента при изготовлении многих лекарственных препаратов. Уксуснокислые бактерии могут наносить вред в виноделии и пивоваренной промышленности, вызывая прокисание вина и пива. Молочнокислое брожение — широко распространенное биохимическое явление, давно известное на примере скисания молока. Под влиянием молочнокислых бактерий (семействоLactobacillaceae)лактоза расщепляется на составляющие ее гексозы — глюкозу и галактозу, которые затем специфическими ферментами превращаются в молочную кислоту. Свертывание молока происходит вследствие того, что молочная кислота отщепляет кальций от казеина, белок превращается в параказеин и выпадает в осадок. Молочнокислые бактерии широко распространены в природе. Они обнаруживаются в молоке, воздухе, на коже, шерсти, в тонком и толстом кишечнике и представлены большим количеством видов палочковидных и кокковидных бактерий, различающихся не только по морфологии, но и физиологическим свойствам (по использованию различных источников углерода и азота). Маслянокислое брожение также широко встречается в природе. Возбудитель маслянокислого брожения был открыт Л. Пастером. На примере маслянокислого брожения Л. Пастер разработал учение об анаэробах. Типичный представитель бактерий маслянокислого брожения — азотфиксирующий Clostridium pasteurianum. Маслянокислые бактерии в больших количествах встречаются в почве, навозе, на растениях, в молоке, сыре. Многие из них являются анаэробами и относятся к роду Clostridium. Маслянокислое брожение — сложный биохимический процесс расщепления углеводов, в ряде случаев жиров и белков, на масляную кислоту, углекислоту и воду, при этом образуется много побочных продуктов — уксусная, молочная, пропионовая и другие кислоты. Из числа других форм брожения чрезвычайно важным является брожение целлюлозы (клетчатки), в которой заложены огромные запасы углерода. Разложение целлюлозы, которая в количественном отношении представляет один из основных компонентов растительных тканей, осуществляется главным образом высоко специализированными в отношении питания аэробными и анаэробными микроорганизмами. Среди аэробных бактерий, расщепляющих целлюлозу, наиболее важны скользящие бактерии рода Cytjphaga. Целлюлоза — единственное вещество, которое они могут использовать в качестве источника углерода. Цитофаги быстро растворяют и окисляют целлюлозу. |
Последнее изменение этой страницы: 2019-04-11; Просмотров: 200; Нарушение авторского права страницы