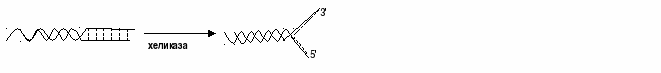|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
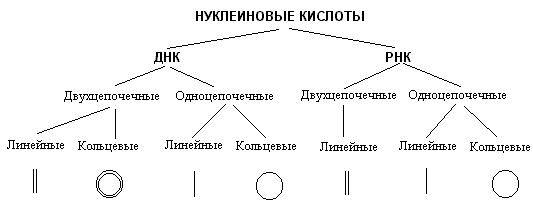
Строение нуклеиновых кислот.Стр 1 из 28Следующая ⇒
Виды РНК. Перечислите их функции. Химический состав нуклеиновых кислот Состав нуклеиновых кислот Нуклеиновые кислоты представляют собой полимеры, построенные из нуклеотидов, соединенных между собой фосфодиэфирными связями. Каждый нуклеотид состоит из остатков азотистого основания, пентозы и фосфорной кислоты. Различают пиримидиновые и пуриновые основания, называемые также соответственно пиримидины и пурины. Пиримидиновые основания являются производными пиримидина:
пуриновые основания – производными пурина: К пиримидинам относятся урацил, тимин и цитозин, к пуринам – аденин и гуанин: В состав ДНК входят тимин, цитозин, аденин и гуанин, в состав РНК – те же основания, только вместо тимина входит урацил. Кроме азотистых оснований, нуклеиновые кислоты содержат пентозы: ДНК – D-дезоксирибозу, а РНК – D-рибозу. Углеводы находятся в виде b-аномера фуранозной формы:
Азотистое основание связывается с углеводом за счет гликозидного гидроксила. Образуется нуклеозид. Схематически образование нуклеозида можно изобразить так:
В состав нуклеиновых кислот входят 8 нуклеозидов, 4 – в состав РНК и 4 – в состав ДНК (рис. 39). Нуклеозиды, входящие в состав РНК:
Нуклеозиды, входящие в состав ДНК:
Рис. 39. Нуклеозиды
Нуклеозид, связанный с остатком фосфорной кислоты, называется нуклеотидом:
При этом остаток фосфорной кислоты может быть связан с 3’- или 5’- атомом углерода:
Сокращенно аденозин-5’-монофосфат обозначается как АМФ. Если нуклеотид образован дезоксорибозой, аденином и одним остатком фосфорной кислоты, то он будет носить название дезоксиаденозинмонофосфат, или сокращенно дАМФ. В таблице 5 представлена номенклатура нуклеотидов. Репликация ДНК Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа: 1. инициация репликации 2. элонгация 3. терминация репликации. Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятиерепликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий[1]. Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окружённый более протяжёнными участками нереплицированной ДНК[1]. В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот[3]. Молекулярный механизм репликации Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у эукариот осуществляется несколькими разными ДНК-полимеразами. Далее происходит закручивание синтезированных молекул по принципусуперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный. Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле. Рис. 6.1. Репликация ДНК
Олигонуклеотид, синтезированный ДНК-полимеразой a и образующий небольшой двухцепочечный фрагмент с матрицей, позволяет присоединиться ДНК-полимеразе d и продолжить синтез новой цепи в направлении 5¢®3¢ по ходу раскручивания репликативной вилки. Выбор ДНК-полимеразой очередного нуклеотида определяется матрицей: включение нуклеотида в синтезируемую цепь ДНК невозможно без предварительного связывания азотистого основания водородными связями с комплементарным нуклеотидом матричной цепи.
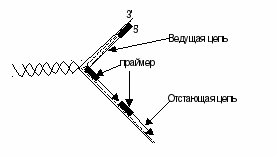
В каждой репликативной вилке идет одновременно синтез двух дочерних цепей. Направление синтеза цепи ДНК совпадает с направлением движения репликативной вилки лишь для одной из вновь синтезируемых цепей (лидирующая цепь). На второй матричной цепи синтез новой цепи осуществляется двумя ферментами: ДНК-полимеразой a и ДНК-полимеразой eв направлении 5¢®3¢, но противдвижения репликативной вилки. Поэтому вторая цепь синтезируется прерывисто, короткими фрагментами, которые по имени открывшего их исследователя называют «фрагменты Оказаки».Дочернюю цепь, синтез которой происходит фрагментами, а потому отстает, называют отстающей цепью. Каждый фрагмент Оказаки содержит праймер. Праймеры удаляет ДНК-полимераза b,после чего присоединяет к ОН-группе на 3¢-конце предыдущего фрагмента дезоксирибонуклеотиды в количестве, равном вырезанному фрагменту и таким образом заполняет брешь, возникающую при удалении рибонуклеотидов. ФерментДНК-лигазакатализирует образование фосфодиэфирной связи между 3¢-ОН-группой дезоксирибозы одного фрагмента и 5¢-фосфатом следующего. Реакция протекает с затратой энергии АТФ. Таким образом из множества фрагментов Оказаки образуется непрерывная цепь ДНК. Терминация синтеза ДНК наступает вследствие исчерпания матрицы при встрече двух репликативных вилок. После окончания репликации происходит метилирование вновь образованных цепей ДНК. Наличие СН3-групп необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов. На каждом конце хромосомы имеются неинформативные повторяющиеся последовательности нуклеотидов – теломеры.В соматических клетках с каждым актом репликации теломеры укорачиваются из-за невозможности достроить ДНК на месте 5¢-праймера. Это укорочение является важным фактором, определяющим продолжительность жизни клетки. Однако в эмбриональных и других быстро делящихся клетках потери концов хромосом недопустимы, так как укорочение хромосом будет происходить очень быстро. У эукариотических клетках имеется фермент теломераза, обеспечивающий восстановление недореплицированных 5¢-концов. В большинстве клеток теломераза неактивна, так как соматическая клетка имеет длину теломерной ДНК, достаточную для времени жизни клетки и её потомства. Небольшая активность теломеразы обнаруживается в клетках с высокой скоростью обновления, таких как лимфоциты, стволовые клетки костного мозга, клетки эпителия и т.д.
С труктура и функции белков Белки (протеины) составляют 50% от сухой массы живых организмов.
Белки состоят из аминокислот. У каждой аминокислоты есть аминогруппа и кислотная (карбоксильная) группа, при взаимодействии которых получается пептидная связь, поэтому белки еще называют полипептидами. Структуры белка Первичная – цепочка из аминокислот, связанных пептидной связью (сильной, ковалентной). Чередуя 20 аминокислот в разном порядке, можно получать миллионы разных белков. Если поменять в цепочке хотя бы одну аминокислоту, строение и функции белка изменятся, поэтому первичная структура считается самой главной в белке.
Вторичная – спираль. Удерживается водородными связями (слабыми).
Третичная – глобула (шарик). Четыре типа связей: дисульфидная (серный мостик) сильная, остальные три (ионные, гидрофобные, водородные) – слабые. Форма глобулы у каждого белка своя, от нее зависят функции. При денатурации форма глобулы меняется, и это сказывается на работе белка.
Четвертичная – имеется не у всех белков. Состоит из нескольких глобул, соединенных между собой теми же связями, что и в третичной структуре. (Например, гемоглобин.) Функции белков: 1) защитная (интерферон усиленно синтезируется в организме при вирусной инфекции); 2) структурная (коллаген входит в состав тканей, участвует в образовании рубца); 3) двигательная (миозин участвует в сокращении мышц); 4) запасная (альбумины яйца); 5) транспортная (гемоглобин эритроцитов переносит питательные вещества и продукты обмена); 6) рецепторная (белки-рецепторы обеспечивают узнавание клеткой веществ и других клеток); 7) регуляторная (регуляторные белки определяют активность генов); 8) белки-гормоны участвуют в гуморальной регуляции (инсулин регулирует уровень сахара в крови); 9) белки-ферменты катализируют все химические реакции в организме; 10) энергетическая (при распаде 1 г белка выделяется 17 кдж энергии). Биосинтез белка Основываясь на данных биспиральной антипараллельной структуры и химического состава ДНК (см. Химия нуклеиновых кислот), а также на данных о значении "активированной" формы энергии для биосинтеза полимерных молекул, А. Корнберг еще в 1955 г. впервые указал на возможность синтеза энзиматическим путем ДНК в бесклеточной системе в присутствии изолированной из Е. coli ДНК-полимеразы и предшественников дезоксирибонуклеозидтрифосфатов; реакция сводится к синтезу новой молекулы ДНК:
Сущность реакции сводится к присоединению мононуклеотидных единиц к свободному 3'-гидроксильному концу ДНК и образованию новой полинуклеотидной цепи в направлении 5'-> 3'; схематически она может быть представлена в виде:
Реакция требует присутствия одноцепочечной ДНК или в крайнем случае небольших фрагментов полинуклеотидов. Пока еще нет полной ясности о значении предобразованной ДНК в действии ДНК-полимераз. Более вероятно мнение, что ДНК служит матрицей, на которой фермент комплементарно и антипараллельно синтезирует новую цепь ДНК. Это можно представить в виде схемы:
Как видно из схемы, последовательность вновь синтезируемого полинуклеотида отражает первичную структуру, и обе цепи по отношению друг к другу являются антипараллельными. Следует указать, что ДНК-полимераза I оказалась наиболее активной в тех случаях, когда добавленная в качестве матрицы молекула ДНК была предварительно денатурирована. В этих случаях соблюдалась не только точная последовательность оснований, но и ряд физических свойств исходной ДНК. Напротив, если в опытах in vitro была добавлена нативная двухцепочечная ДНК, то вновь синтезированный полинуклеотид значительно отличался от ДНК и, в частности, не обладал свойственной ДНК биологической активностью.
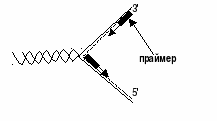
Были предприняты другие подходы к выяснению механизма полимеразной реакции. В лаборатории Корнберга был открыт фаг (ФХ174), содержащий одноцепочечную кольцевую ДНК; когда эта молекула была использована в качестве матрицы в ДНК-полимеразной реакции, то в этих условиях была получена биологически активная ДНК фага. В этой работе важную роль сыграл другой фермент, названный ДНК-лигазой, обладающий способностью катализировать репарацию ДНК, соединение концов разрывов в молекуле ДНК. Было показано, что в процессе репликации одноцепочечная ДНК фага ФХ174 проходит стадию образования двухцепочечной кольцевой ДНК. Применив ряд остроумных подходов, Корнберг и сотр. в опытах in vitro создали искусственную молекулу фага ФХ174, обладающую способностью поражать (инфицировать) Е. coli, вызывая лизис бактерии. Последовательность событий в этой работе может быть представлена в следующей схеме, где исходную молекулу кольцевой ДНК фага ФХ174 пометим плюсом (+), а вновь синтезируемую молекулу-минусом (-) (рис. 106). В исследованиях Меселсона и Сталя получила подтверждение гипотеза полуконсервативного механизма синтеза ДНК в клетках, означающего наличие в каждой из двух дочерних молекул одной из родительских цепей ДНК (рис. 107). Репликация ДНК, как правило, начинается с какой-нибудь одной точки на обеих цепях, и направление роста каждой цепи также идет в одном направлении. Кроме того, как было указано выше, ДНК-полимераза I не реплицирует нативную двухцепочечную ДНК и осуществляет синтез только в одном направлении, а именно в направлении 5'->3'. Была предложена интересная гипотеза, объясняющая репликацию обеих цепей нативной молекулы ДНК. Этот механизм требует наличия трех ферментов: ДНК-полимеразы, ДНК-лигазы и эндонуклеазы. Репликация начинается с разрыва в одной из двух цепей под действием эндонуклеазы. Затем к этому месту присоединяется ДНК-полимераза и начинается непрерывный синтез нового олигонуклеотида на одной из двух родительских цепей в направлении 5'->3', обратный направлению исходной цепи ДНК. Из второй цепи родительской ДНК идет синтез прерывистый, сопровождающийся образованием фрагментов ДНК, также в направлении 5'->3' с последующим объединением фрагментов ДНК при участии ДНК-лигаз в единую полинуклеотидную молекулу. Подобный механизм челночного синтеза ДНК легко объясняет фактические данные накопления коротких фрагментов ДНК у Е. coli во время репликации ДНК. Совсем недавно было показано, что инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на материнской ДНК необычного затравочного олигорибонуклеотида, названного праймером, со свободной гидроксильной группой, у 3'-углеродного атома рибозы. Этот олигорибонуклеотид, содержащий около 50 нуклеотидных остатков, синтезируется комплементарно на матрице ДНК при участии особого фермента - РНК-полимеразы (см. ниже). Предполагается, что именно с этой точки концевого 3'-гидроксила рибозы праймера начинается истинный синтез дочерней цепи ДНК, комплементарной родительской. В дальнейшем этот фрагмент РНК, ковалентно присоединенный к новообразованной цепи ДНК, разрушается под действием нуклеаз и возникшая брешь застраивается олигодезоксирибонуклеотидом при помощи той же ДНК-полимеразы. Вполне допустимо предположение, что синтез праймера изолигорибонуклеотида имеет биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
Классификация белков Функции белков Классификация по функциям По выполняемым функциям белки можно разделить на структурные, питательные и запасные белки, сократительные, транспортные, каталитические, защитные, рецепторные, регуляторные и др.
Структурные белки К структурным белкам относятся коллаген, эластин, кератин, фиброин. Белки принимают участие в формировании клеточных мембран, в частности, могут образовывать в них каналы или выполнять другие функции Питательные и запасные белки Питательным белком является казеин, основная функция которого заключается в обеспечении растущего организма аминокислотами, фосфором и кальцием. К запасным белкам относятся яичный белок, белки семян растений. Эти белки потребляются во время развития зародышей. В организме человека и животных белки в запас не откладываются, они должны систематически поступать с пищей, в противном случае может развиться дистрофия.
Сократительные белки Сократительные белки обеспечивают работу мышц, движение жгутиков и ресничек у простейших, изменение формы клеток, перемещение органелл внутри клетки. Такими белками являются миозин и актин. Эти белки присутствуют не только в мышечных клетках, их можно обнаружить в клетках практически любой ткани животных.
Транспортные белки Гемоглобин, рассмотренный в начале параграфа, является классическим примером транспортного белка. В крови присутствуют и другие белки, обеспечивающие транспорт липидов, гормонов и иных веществ. В клеточных мембранах находятся белки, способные переносить через мембрану глюкозу, аминокислоты, ионы и некоторые другие вещества. На рис. 24 схематически показана работа переносчика глюкозы. Белки-ферменты Каталитические белки, или ферменты, представляют собой самую многообразную группу белков. Почти все химические реакции, протекающие в организме, протекают при участии ферментов. К настоящему времени открыто несколько тысяч ферментов. Более подробно они будут рассмотрены в следующих параграфах.
Защитные белки К этой группе относятся белки, защищающие организм от вторжения других организмов или предохраняющие его от повреждений. Иммуноглобулины, или антитела, способны распознавать проникшие в организм бактерии, вирусы или чужеродные белки, связываться с ними и способствовать их обезвреживанию. Другие компоненты крови, тромбин и фибриноген, играют важную роль в процессе свертывания крови. Они предохраняют организм от потери крови при повреждении сосудов. Под действием тромбина от молекул фибриногена отщепляются фрагменты полипептидной цепи, в результате этого образуется фибрин: фибриноген Образовавшиеся молекулы фибрина агрегируют, формируя длинные нерастворимые цепи. Сгусток крови вначале является рыхлым, затем он стабилизируется за счет межцепочечных сшивок. Всего в процессе свертывания крови участвует около 20 белков. Нарушения в структуре их генов является причиной такого заболевания, как гемофилия – сниженная свертываемость крови.
Рецепторные белки Клеточная мембрана является препятствием для многих молекул, в том числе и для молекул, предназначенных для передачи сигнала внутрь клеток. Тем не менее клетка способна получать сигналы извне благодаря наличию на ее поверхности специальных рецепторов, многие из которых являются белками. Сигнальная молекула, например, гормон, взаимодействуя с рецептором, образует гормон-рецепторный комплекс, сигнал от которого передается далее, как правило, на белковый посредник. Последний запускает серию химических реакций, результатом которых является биологический ответ клетки на воздействие внешнего сигнала.
Регуляторные белки Белки, участвующие в управлении биологическими процессами, относят к регуляторным белкам. К ним принадлежат некоторые гормоны. Инсулин и глюкагон регулируют уровень глюкозы в крови. Гормон роста, определяющий размеры тела, и паратиреоидный гормон, регулирующий обмен фосфатов и ионов кальция, являются регуляторными белками. К этому классу белков принадлежат и другие протеины, участвующие в регуляции обмена веществ. Б. Метилирование про-иРНК. Чаще всего происходит у бактерий, у которых имеется специальный аппарат защиты от чужеродной ДНК (вирусной, фаговой). Этот аппарат состоит из целого ряда ферментов разрезающих чужеродную ДНК или РНК в определённых сайтах в которой находится специфическая последовательность нуклеотидов. Ферменты носят название – рестриктазы. Понятно, что собственный, только что синтезированный РНК-транскрипт, также может быть подвергнут атаке рестриктаз. Чтобы это не случилось специальные ферменты, называемые метилазы,метилируют собственный РНК-транскрипт в тех сайтах, которые могут быть разрезаны собственными ферментами. У эукариот РНК-транскрипт метилируется в меньшей степени. В. Кэпирование 5’конца. Заключается в химическом и конформационном изменении 5’конца синтезированной РНК. Кэпирование происходит в момент синтеза РНК, ещё до её отделения. Процесс заключается в присоединении к свободному концу про-РНК специальных химических веществ, которые изменяют конформацию концевого участка. Кэпирование необходимо для инициации процесса трансляции. Специальные ферменты присоединяют к 5’концу про-иРНК ГДФ (гуанозиндифосфат), а затем метилируют его. 1. Инициирует синтез белка. 2. Предохраняет про-иРНК от распада. 3. Участвует в удалении интронов. Г. Полиаденилирование. Это процесс присоединения к 3’ концу про-иРНК 100 – 200 остатков адениловой кислоты. Эти остатки носят название поли-А последовательности (поли-А хвосты). Полиаденилированию подвергаются не все про-иРНК. Например, молекулы всех типов гистонов не содержат поли-А последовательности. Полиаденилирование предохраняет иРНК от разрушения. На растущей цепи и-РНК имеется специальная последовательность нуклеотидов (ААУААА). Особый фермент ( полиА-полимераза) находит это сочетание нуклеотидов, разрезает про-иРНК в этом месте и формирует полиадениловый хвостик. Значение поли –А последовательностей: 1. Облегчают выход иРНК из ядра в цитоплазму. 2. Предохраняют иРНК от разрушения. Недавно было выявлено ещё одно интересное свойство поли-А последовательностей – они участвуют в терминации синтеза про-иРНК. РНК-полимераза, формируя последовательность ААУААА в про-иРНК, получает сигнал о завершении синтеза РНК-транскрипта. Но синтез сразу не прекращается. Полная остановка его наступает после того, как РНК-полимераза встречает на матричной нити ДНК специфическую последовательность нуклеотидов (у разных генов она разная), которая и даёт окончательный сигнал о прекращении синтеза РНК. Д. Сплайсинг. В РНК-транскрипте содержится определённое количество нуклеотидных последовательностей, которые были необходимы для успешного завершения трансляции и последующей модификации транскрипта (кэпирования, полиаденилирования и т.д.). Для выполнения основной роли РНК в цитоплазме – трансляции, эти последовательности не только не будут иметь функционального значения, но могут помешать нормальному течению синтеза белка. Поэтому в клетке предусмотрен механизм освобождения первичного транскрипта от целого ряда последовательностей, не имеющих решающего значения в трансляции. К таким последовательностям прежде всего относят интроны. Ген, с которого транскрибировалась про-иРНК содержит кодирующие и некодирующие последовательности. Кодирующие последовательности гена определяют аминокислоту и их последовательность в белке. Не кодирующие последовательности таким свойством не обладают. Кодирующие и некодирующие последовательности в гене чередуются, и их количество зависит от индивидуальных генов. В первичном транскрипте также содержатся кодирующие и некодирующие последовательности. Такая организация генов и про-РНК характерна для эукариот. Некодирующие последовательности про-иРНК носят название интроны, а кодирующие –экзоны. Длина интронов может быть от 50 до 12000 нуклеотидов. Ген начинается и кончается экзоном. Прерывистое строение гена характерно для большинства эукариот. Интроны могут содержать все виды РНК – иРНК, тРНК, рРНК. Вся совокупность экзонов (кодирующих белки) в геноме человека занимают всего 1,1 – 1,4 %. Средний ген человека содержит 9 интронов. По мере упрощения организации организмов совокупная величина их экзонов возрастает (например у бактерий она равна 86%). В вырезании интронов из РНК-транскрипта и сшивании оставшихся экзонов, принимает участие многокомпонентный комплекс. Основными его составляющими являются малые ядерные РНК (мяРНК) и белки-ферменты. В целом комплекс носит название малые ядерные рибонуклеопротеиды, мяРНП или сплайосома. Сам процесс достаточно сложен и состоит из нескольких этапов (см. рис. 58). 1. Формирование сплайосомы. К началу и концу интрона прикрепляются фрагменты белка и мяРНК (рис. 56, Д) формируя сплайосому. (рис. 56, Д) Прикрепление комплекса мяРНП (рис. 56, Е). Рис. 61. Схема сплайсинга (объяснение в тексте). 2. Сближение соседних экзонов, за счёт образования петли интрона. Разрезание на границе экзон-интрон и соединение соседних (первого и второго) экзонов(рис. 56, В). 3. Удаление и разрушение петли и сплайосоме (рис. 56, Г, Ж). Необходимо отметить, что при повреждении (мутации) интрона сплайсинг может быть не закончен, интрон не вырезан и конченый продукт – иРНК будет нести несвойственные ей последовательности нуклеотидов. Понятно, что это может привести к нарушению трансляции и выключению из метаболизма определённого белка МЗ. Процессинг (или созревание РНК) это процесс превращения только что синтезированной, не активной РНК (про-иРНК) в функционально активную РНК. Процесс связан со структурными и химическими модификациями про-иРНК. Происходит в ядре до момента выхода РНК в цитоплазму. Состоит из нескольких этапов: присоединение про-иРНК к белку, метилирование некоторых оснований, маркировка одного из концов, полиаденилирование другого (противоположного) конца, вырезания интронов и сшивание экзонов. Последние два процесса носят название сплайсинг. Метилирование ДНК После завершения репликации происходит метилирование нуклеотидных остатков вновь образованных цепей ДНК. Метальные группы присоединяются ко всем остаткам аденина в последовательности - GATC -. Количество метилированных оснований равно примерно 1-8%. Модификация происходит при участии ферментов, использующих в качестве источника метальных групп S-аденозилметионин (SAM). Присоединение метальных групп к остаткам аденина и цитозина не нарушает комплементарности цепей (рис. 4-18).
Наличие метальных групп в цепях ДНК необходимо для формирования структуры хромосом, а также для регуляции транскрипции генов.
Рис. 4-18. Метилирование остатков аденина в последовательности - GATC -. В течение нескольких минут после репликации, пока не произошло метилирование, новая цепь ДНК отличается от матричной цепи. Метилирование ( methylation) - это химическая модификация, катализируемая ферментом, реакция добавления метильных группп (CH3) на специфические сайты белков, ДНК и РНК. Одна из форм метилирования, наиболее распространенная у млекопитающих, представляет собой превращение цитозина в 5-метилцитозин в последовательности CpG . Метилирование может предотвращать расщепление ДНК в сайте узнавания рестрикционного фермента . Например, рестриктаза Hpall расщепляет CCGG , но не Cm5CGG . Реакция ДНК-метилирования катализируется ферментом ДНК-метилтрансферазой, который осуществляет перенос метильной группы с S-аденозилметионина на цитозин, стоящий перед гуанином. У человека и большинства млекопитающих ДНК-метилирование - естественная модификация ДНК, и воздействует только на основание цитозин (С), стоящий перед гуанином (G), т.е. метилирование происходит только в CpG-динуклеотидах. У растений 5-метилцитозин можно обнаружить в динуклеотидах CG и тринуклеотидах CNG (Т-С, A или Т). 70-80% всех CpG-динуклеотидов в человеческом геноме метилированы. Однако в нормальной ткани метилирование происходит, прежде всего, в областях, где плотность CpG динуклеотидов низка, и большинство CpG - островков в норме полностью неметилированы. Но есть исключения из этого правила: метилированные промоторные области генов на инактивированной Х хромосоме, например, ген FMR1 (Li et al 1993 , Singer-Sam et al. 1993 ) и импринтированные гены. Так экспрессия таких генов как IGF2 , SNRPN , Znf-21 , Par-5 идет только с отцовской хромосомы, а WT1 , Mash2 , p57 - только с материнской. Также профиль метилирования может меняться в течение жизни организма. Некоторые гены, экспрессирующиеся в эмбриональном периоде, перестают функционировать к моменту рождения особи. А для таких генов как ген E-CAD и ген DBCCR1 показано метилирование в нормальных тканях по мере старения организма. Функция ДНК метилирования до сих пор не полностью ясна. За последние годы было предложено несколько версий роли ДНК метилирования: контроль экспрессии гена, контроль целостности хромосомы, контроль пререкомбинантных событий. Гипотезой о роли ДНК-метилирования в геноме является предположение о том, что это защитный механизм против встраивания в геном "паразитных" последовательностей типа ретровирусных элементов ( Issa, 1994 ). К структурам, обеспечивающим этот процесс, относятся: несколько ДНК-метилтансфераз, деметилазы, центры метилирования, которые инициируют метилирование, и вероятно, связаны с необычными третичными структурами ДНК. Эмбрионы мышей, несущие нонсенс мутацию в гене метилтрансферазы, не развиваются дальше 8-го сомита, но при этом наблюдается низкий, но постоянный уровень метилцитозина. Это может говорить о том, что имеются и другие гены, обеспечивающие метилирование. Метилированные основания в ДНК обнаружены свыше 50 лет назад [ Hotchkiss, ea 1948 ]. ДНК прокариот содержит модифицированные основания N6-метиладенин и 5-метилцитозин , тогда как ДНК высших эукариот - в основном 5-метилцитозин [ Wyatt, ea 1951 , Shapiro, ea 1960 , Doscocil, ea 1965 , Vanyushin, ea 1970 , Vanyushin, ea 1968 ]. Метилирование остатков цитозина ДНК имеет место у бактерий, растений, животных, в том числе млекопитающих (включая человека), но отсутствует у дрожжей, нематод и дрозофилы [ Мазин ea 1984 , Мазин ea 1989 , Doerfler, ea 1983 , Bird, ea 1995 ].(Обнаружено, что в ДНК дрозофилы содержится 5-метилцитозин ( Growher, Н., Leismann. О.,, 2000 ) Метилирование осуществляется ферментативно в первые минуты после репликации ДНК, т.е. пострепликативно. Поскольку нуклеотидная последовательность ДНК при этом не меняется, метилирование по сути своей - событие эпигенетическое [ Baylin, ea 1998 ]. Оно, хотя и является стабильной и наследуемой модификацией, в принципе обратимо под воздействием деметилирующих агентов или ферментов и тем самым принципиально отличается от мутаций ДНК. В общебиологическом плане феномен метилирования является элементом системы распознавания "свой-чужой". Благодаря существующей в бактериях системе метилирования-рекогниции ( рестрикции-модификации ) клетки способны идентифицировать свой генетический материал и отличать его от инородных молекул, проникших в клетку тем или иным способом. Уничтожение последних позволяет поддерживать генетическую стабильность вида. Иногда один фермент имеет две активности - метилазную и эндонуклазную: в большинстве случаев определенный сайт ДНК распознается двумя ферментами, один из которых метилирует его, а другой расщепляет. В целом система функционирует таким образом, что метилазы "метят" специфические последовательности собственной ДНК, а чувствительные к метилированию рестрикционные эндонуклеазы узнают и расщепляют те из последовательностей. которые соответствующей метки не имеют. Так, бактериальная клетка защищает себя от вторжения чужеродных молекул. Например, ДНК проникшего в бактериальную клетку фага расщепляется в определенных сайтах специфическими эндонуклеазами, в то время как те же последовательности в собственной ДНК защищены от расщепления, поскольку метилированы [ Noyer-Weidner, ea 1993 ]. Роль метилирования ДНК как компонента клеточной "иммунной системы", предназначенной для уничтожения чужой или излишней ДНК (или подавления ее функций), сохраняется по-видимому, на протяжении эволюции, но конкретные механизмы реализации этой задачи могут быть существенно иными [ Bestor, ea 1990 ]. Например, клетки Neurospora элиминируют нежелательные повторяющиеся последовательности посредством интенсивного их метилирования, за которым следует накопление точковых мутаций из-за высокой мутабельности остатков 5-метилцитозина [ Selker, ea 1993 ]. Подобным же образом в клетках грызунов и человека интегрированные вирусные последовательности могут подвергаться метилированию и обусловленному им стабильному блоку транскрипции [ Doerfler, ea 1993 ]. Известно также, что инактивации тем же способом нередко подвергаются трансгены у мышей [ Sasaki, ea 1993 ]. В широком эволюционном плане переходы от прокариот к эукариотам и от беспозвоночных к позвоночным сопровождались, по-видимому, резким увеличением числа генов [ Bird, ea 1995]. Эти драматические изменения вызвали к жизни, видимо, и новые способы ограничения нежелательной активности "лишних" генов - формирование ядерной мембраны и нуклеосомную организацию хроматина в первом случае, функциональную переориентацию системы метилирования - во втором. Если у беспозвоночных она сводилась к подавлению активности потенциально опасных последовательностей ДНК (таких как вирусы и транспозоны), то у позвоночных ее назначение - еще и стабильная репрессия эндогенных генов (гены инактивированной хромосомы X, импринтированные гены, часть тканеспецифичных генов). Профиль метилирования, сильно влияющий на функциональное состояние гена, стабильно передается в ряду клеточных поколений. С этой точки зрения, для организмов с большой продолжительностью жизни и интенсивной тканевой регенерацией (позвоночные, растения) надежная система эпигенетической наследственности (типа метилирования ДНК) жизненно необходима. В противоположность этому у маленьких животных и животных с малой продолжительностью жизни, т.е. в ситуациях, когда значительное новообразование клеток отсутствует. такой необходимости нет. Предполагают, что именно этим обстоятельством объясняется отсутствие системы метилирования ДНК у нематод и дрозофилы [ Jablonka. ea 1995 ]. Рестрикционный анализ – классический метод анали- за метилирования, основанный на неспособности некото- рых рестрикционных ферментов разрезать метилированные последовательности ДНК. Пара ферментов HpaII–MspI рас- познает последовательность CCGG, но рестриктаза HpaII не способна разрезать ДНК, в которой метилирован вну- тренний С, что позволяет использовать эти ферменты для быстрого определения метилированных участков. Рестрик- ционный метод имеет несколько недостатков. Главный – не все CG расположены в последовательностях CCGG, что может привести к появлению ложноотрицательных резуль- татов. Для проведения рестрикции и последующего анализа необходимо не менее 0,5–1 мкг ДНК [26]. Полученные после рестрикции фрагменты можно анали- зировать несколькими способами: масс-спектрометрический анализ, саузерн-блотт, лигирование с мечеными нуклеотида- ми, полимеразно-цепная реакция (ПЦР) и др. 1.1. Масс-спектрометрический анализ. Для анализа ме- тилирования ДНК обычно используют специфическую ре стрикцию совместно с MALDI-TOF-масс-спектрометрией или электроспрей-ионизацией. Основным требованием для осуществления масс-спектрометрического анализа являет- ся высокая химическая чистота исследуемого соединения. Особенности анализатора налагают ограничение на раз- мер исследуемых молекул ДНК: средний предел измерений около 30 кДа (100 bp), нижняя граница не менее 600 Да [2]. Масс-спектрометрический анализ не удобен для применения в клинической практике, но в сочетании с рестрикционным анализом, гибридизацией и другими методами лег в основу высокопроизводительных систем для сканирования метили- рования геномной ДНК [16]. 1.2. Полимеразное лигирование с мечеными нуклеотида- ми. Меченые нуклеотиды (радиоактивные 3 H-dCTP/-dGTP, 32P-dCTP/-dGTP или флюоресцентно-меченые dCTP/dGTP) присоединяют по липким концам фрагментов, которые об- разуются после рестрикции (рис. 3). Метод позволяет опре- делить количество 5mC в образце ДНК. Долю метилирован- ного цитозина (в норме 80–90%) рассчитывают по формуле: [MspI] - [HpaII] ––––––––––––– %, [MspI] где [MspI] и [HpaII] – радиоактивность/флюоресценция об- разца после рестрикции соответствующей рестриктазой. 1.3. Саузерн-блотт позволяет определить общий статус метилирования CpG-островков интересующего региона, но требует довольно большого количества (не менее 5 мкг) геномной ДНК и не обладает большой чувствительностью. Количество гибридизованных фрагментов может быть до- вольно высоким. Для CG-богатых последовательностей, где рестрикционные фрагменты имеют длину 100–500 bp, для фореза применяют 1,2% агарозный гель, который до- вольно сложно использовать для блотта [9]. В то же время клеточные линии и особенно ткани поликлональны и содер- жат смешанные паттерны метилирования, поэтому анализ бандов довольно сложен. Тем не менее этот метод гораздо проще и быстрее, чем бисульфитная модификация [17]. 1.4. ПЦР-анализ – более чувствительный метод, позволя- ющий определить статус метилирования определенного ре гиона, сочетает реакцию рестрикции и ПЦР. После рестрикции проводят ПЦР с праймерами, фланкирующими рестрик- ционные сайты, амплификация проис- ходит только в том случае, если ДНК была метилирована и не подверглась разрезанию. Так же как и Саузерн-блотт, этот метод может определить метилиро- вание только в сайтах рестрикции ме- тилчувствительных ферментов. Более того, при полной рестрикции неметили- рованной ДНК ПЦР не происходит, что может привести к ложноположительно- му результату. Неполная рестрикция и присутствие малого количества метили- рованных аллелей дают сходные резуль- таты, и этот метод нельзя использовать для определения, на- пример, гиперметилирования онкосупрессоров, когда клетки с метилированными аллелями составляют лишь небольшую фракцию в популяции [18]. 2. Бисульфитная модификация ДНК. В одноцепочечной ДНК бисульфит натрия преимущественно деаминирует ци- тозин до урацила (и очень медленно деаминирует 5-метил- цитозин в тимин) (Shapiro et al., 1973). В 1992 г. Frommer и др. предложили использовать различие в реакционности би- сульфита для геномного секвенирования 5-метилцитозино- вых остатков. Проводят полную денатурацию геномной ДНК и обработку бисульфитом в условиях, при которых цитозин стехиометрически конвертируется в урацил (рис. 4), но 5-ме- тилцитозин не подвергается изменениям. Результатом би- сульфитной реакции становятся цепи ДНК, которые больше не комплементарны [18, 19]. Бисульфитная конверсия – очень мощный метод, который используют для определения метилирования ДНК не только простым секвенированием, но и в сочетании с ПЦР, SnuPE, MethyLight, олигонуклеотидными микроматрицами, HPLC, пиросеквенированием и др. [10]. 2.1. Бисульфитная ПЦР. При ПЦР-амплификации ре- гиона интереса в бисульфитобработанной ДНК весь ура- цил, ранее бывший цитозином, и тимин амплифицируются и считываются так же, как тимин, а 5-метилцитозин – как цитозин. После бисульфитной ПЦР ДНК можно клониро- вать и секвенировать – процесс длительный и технически довольно сложный. Без клонирования ПЦР-продуктов ме- тод даже менее чувствителен, чем Саузерн-блотт (должно быть метилировано не менее 25% аллелей) [15]. Метод бисульфитной ПЦР используют также в коммер- ческих наборах для определения метилирования. Quantative MethyLight (Epigenomics): детекцию метилированных и не- метилированных участков производят при помощи динуклео- тидов CG и TG, меченных разными флюорофорами (рис. 5, а) [5, 27]. В наборе HeavyMethyl (Epigenomics) специфиче- ский блокатор препятствует амплификации бисульфитмо- дифицированной ДНК, и амплификация происходит только при отсутствии метилирования. Детекция осуществляется по присоединению флюоресцентно-меченого CG рис. 5, б) [27]. 2.2. Метилспецифичная ПЦР (MSP) позволяет опреде- лить статус метилирования интересующей последователь- ности вне зависимости от количества метилированных CpG и не требует клонирования и использования метилчувстви- тельных рестриктаз. После обработки ДНК бисульфитом проводят амплификацию с праймерами, специфичными для метилированных и неметилированных участков ДНК. ПЦР- праймеры в этом случае нужно подобрать таким образом, чтобы они специфически связывались с только бисульфитмо- дифицированной цепью. Праймеры для каждой цепи будут отличаться в той позиции, где были CG оригинальной после- довательности (см. таблицу). Метод требует небольшого количества геномной ДНК. Отличается высокой чувствительностью (0,1% метилирован- ных аллелей), поэтому представляется весьма удобным для анализа клинических образцов [20]. 2.3. Single Nucleotide Primer Extension (SnuPE) – метод, который можно использовать для точного анализа метили- рования в определенной позиции ДНК. После бисульфитной обработки ДНК проводят отжиг праймеров, которые закан- чиваются непосредственно перед анализируемым сайтом. Затем осуществляют две реакции удлинения цепи – с радио- активным dCTP и радиоактивным dTTP (рис. 6). В этих реак- циях только один нуклеотид добавляется к праймерам. После окончания реакции продукты разгоняются в акриламидном геле, и анализируется радиоактивность меченых праймеров. Для этого метода также можно использовать флюоресцентно- меченые нуклеотиды [21]. 3. Иммунопреципитация ДНК. В этом методе исполь- зуют специфические антитела к белкам, способным се- лективно связывать метилцитозин, – таким как MeCP2 и MBD2 [6]. Метод относительно чувствительный и не требует рестрикции геномной ДНК или бисульфитной обработки. Данные, полученные методом иммунопреци- питации, проще анализировать и интерпретировать, чем, например, после бисульфитной конверсии. В то же время иммунопреципитация позволяет получить данные только об общем метилировании и не применима к изучению кон- кретного гена. 4. Микроматричный анализ метилирования ДНК. Кон- струирование микроматриц (micro-array) для анализа мети- лирования ДНК основывается на трех методах: рестрикции, аффинной очистке и бисульфитной конверсии. Микрома тричная методология позволяет проводить сканирование всей геномной ДНК. Коммерческие микроматрицы NimbleGen ("Roche") вы- пускают в нескольких вариантах, что позволяет проводить анализ метилирования отдельных хромосом человека. Микроматрицы NimbleGen включают все классифициро- ванные CpG-островки, в том числе и редко встречающиеся [22]. Affymetrix (Santa Clara) – несколько типов платформ для анализа метилирования. Они требуют очень малого количества (не более 100 нг) ДНК, что дает возможность использовать эти платформы для анализа метилирова- ния в эмбриогенезе и проводить анализ малых образцов [23].
Ацетилирование В новосинтезированных гистонах ацетилированы К5, К12 гистона Н4 и К18, К1 гистона Н3. В течение транспорта из цитоплазмы в ядро, гистоны приобретают дополнительные модификации (H3K9me, H3K56Ac). Ацетилирование гистонов, как правило, ведет к активации транскрипции генов. В транскрипционно активных областях хроматина высокое содержание моноацетилированных гистонов H3 и H4, что свидетельствует об их особой роли в процессе активации транскрипции. Моноцетилирование К6 – главная модификация гистона H4 в клетках животных. Удаление фермента, катализирующего модификацию H3K16 приводит к фрагментации ядра, что еще раз свидетельствует о важной роли этой модификации. Ацетилированные гистоны располагаются в области промотеров генов. Ацетилирование гистонов происходит быстрее, чем метилирование. Метилирование Метилированные гистоны (за исключением H3K4me) располагаются по всей области генов. Метилирование гистонов играет наиболее значительную роль в создании эпигенетической информации, в то время как остальные модификации имеют модулирующую функцию (например, могут иметь роль сигнальных молекул для приема внешнего сигнала хроматином). Активно транскрибируемые районы генов: H3K4me2,3; H3K36me2,3 Репрессия транскрипции: H3K27me3; H4K20me3 Фосфорилирование Фософорилирование гистонов совпадает с началом митоза, который начинается с фосфорилирования линкерного гистона H1. Хвосты (N-концевые части) коровых гистонов подвергаются фосфорилированию во время деления клетки (Т3, S10, S28 гистона Н3 и S1 гистона Н4). Метилирование ДНК Теперь что представляет собой метилирование ДНК. Это результат процессов, в ходе которых на определённые нуклеотиды в составе ДНК садится или же наоборот удаляется метильная группа. Такая метильная группа может исполнять роль метки-закладки, информирующей белковые комплексы как им себя вести относительно этого участка ДНК. Например, если метильные метки расположены на промоторе (последовательности нуклеотидов ДНК на которую садится РНК-полимераза чтобы начать синтез РНК) гена, то это обычно приводит к ингибированию (репрессии) данного гена. Удаление с промотора метильных групп (деметилирование) приводит к активации (дерепрессии) такого гена. ДНК построена из четырёх нуклеотидов, но у эукариот метилированию обычно подвергается только один из них – цитозин. Посадку метильной группы осуществляет фермент ДНК-метилтрансфераза. Следует при этом отметить, что регуляция посредством метилирования ДНК характерна не для всех организмов. Так, например у плодовой мушки дрозофилы число таких метилированных цитозинов ничтожно мало – всего 0.03%, а у червячка C. elegans и дрожжей и того меньше ([21]). У мышки их заметно больше – порядка 4%. Казалось бы так ли уж важно понимать процессы метилирования ДНК для того чтобы разобраться в механизмах старения, ведь у плодовой мушки и у дрожжей эти процессы не играют существенной роли, однако они стареют также как и другие организмы. Почему важно изучать роль метилирования ДНК в механизмах старения? Начнём с того, что метилирование ДНК как правило, исчезает при образовании зиготы – клетки, образующейся в результате оплодотворения и с которой начинается процесс развития нового организма. Затем, по мере развития организма, происходит постепенное накопление всё новых и новых метильных групп. Причем характер метилирования, регулирующего генную активность (экспрессию) при делении клеток передается вновь образованным клеткам. Считается, что метилирование ДНК позволяет стабилизировать экспрессию генов в клетках, закрепляя «клеточную память» в клетках, которые делятся и дифференцируются из эмбриональных стволовых клеток в клетки различных специализированных тканей организма. Поскольку изменение метилирования ДНК закреплено клеточной памятью и стабильно наследуется клетками-потомками данной клетки, но не является само по себе мутацией ДНК, его можно назвать эпигенетическим. ([22]) Все соматические клетки организма (за исключением иммунных клеток вроде T и B клеток, а также эритроцитов, которые при созревании вообще теряют ядро), несмотря на значительные различия в их физиологических функциях, имеют ДНК с одной и той же последовательностью нуклеотидов, но отличающуюся по характеру метилирования. Этот феномен — что в разных клетках одного и того же организма ДНК метилирована по-разному, Ванюшин с соавторами обнаружили ещё в 1970 м году, что уже тогда позволило им первыми заявить, что метилирование ДНК – механизм регуляции экспрессии генов и клеточной дифференцировки ([23]). Было обнаружено что после рождения в ходе старения у коровы, крысы и у мышей понижается содержание метилированного цитозина, причем понижение это происходит главным образом на участках ДНК, несущих многократно повторяющиеся последовательности ([24] [25]). При этом была подмечена закономерность — Mus musculus теряет участки метилирования со скоростью порядка 4,7 Х 104 (в месяц примерно 0.012% от уровня новорожденного), тогда как Peromyscus leucopus теряет 5-метилцитидин со скоростью 2.3 Х 104 (в месяц примерно 0.006% от уровня новорожденного) и при этом живет в два раза дольше чем Mus musculus. У человека общий уровень метилирования ДНК в соматических клетках также неуклонно снижается. В фибробластах человека при приближении репликативного старения, когда способность клетки к делениям заметно падает, происходит общее снижение уровня метилирования ДНК (главным образом в районах бедных на содержание генов, поздно реплицирующихся участков, а также LAD (lamina-associated domain) – доменах ДНК примыкающих к ядерной мембране (что сопровождает отслаивание гетерохроматина от ядерной ламины и замену ламина А на его укороченный аналог – прогерин ([26]) ), Все эти участки обычно несут метки гистонов неактивного хроматина — гетерохроматина). Вместе с тем происходит очаговое гиперметилирование там, где расположены CpGi островки, локализованные на генных промоторах, где метилирование ДНК приводит к ингибированию генов. По мнению авторов этого исследования и их последователей, постаревшие клетки очень похожи на раковый эпигеном ([27] [28]).
По статье ( ) Эпигенетические изменения при старении гемопоэтических стволовых клеток, приводят к увеличению числа случаев возрастных миелопролиферативных заболеваний и злокачественных опухолей миелоидного типа, а также к повышенному риску тромбоза сосудов и снижению сопротивляемости организма к инфекции. В отличие от других клеток организма метилирование ДНК при старении гемопоэтических стволовых клеток не понижается, а наоборот повышается Однако в стволовых клетках предшественниках клеток крови, которые сохраняют способность к делению до глубокой старости организма метилирование ДНК не снижается, а перераспределяется: усиливается в участках генома связанных с лимфоидной и эритроидной специализацией. В результате при старении количество образуемых клеток этих типов снижается ([29]). CpG участки и CpG островки
Участок ДНК с CpG динуклеотидом (серыми столбиками обозначены водородные связи с противоположной комплементарной цепью ДНК). Фиолетовая стрелка указывает на метильную группу 5-метилцитозина. Метилирование ДНК в CpG динуклеотидах обычно симметрично. Обычно метилирование, связанное с изменениями активности генов, происходит на цитозине расположенном на так называемых CpG участках ДНК, где за нуклеотидом цитозина в направлении от 5I (пять штрих) к 3I(три штрих) (исходя из нумерации атомов углерода в молекуле сахара пентозы, см. рисунок) следует нуклеотид гуанина (это часто обозначают так: 5I → 3I). С развитием технологии секвенирования накопились данные о том что помимо метилирования на CpG участках существует также достаточно значимое не-CpG-метилирование, когда в окружении 5-метилцитозина вместо гуанина стоят другие нуклеотиды. Не-CpG метилирование имеет несколько свойств, которые отличают его от CpG метилирования. Так, в отличие от метилирования участков CpG, которое обычно симметрично, то есть метилирован цитозин на обеих цепях ДНК (гуанину комплементарно противостоит цитозин в противоположной цепи), при не-CpG-метилировании метильная группа чаще сидит только на одной из цепей ДНК — то есть не симметрично ([31]). Скопления таких CpG динуклеотидов называют CpG островками и обозначают CpGi. Такие CpGi можно встретить более чем у половины всех генов составляющих геном организма. Длина таких островков обычно не превышает тысячи нуклеотидов. CpG динуклеотиды островков CpGi составляют только 1-2% всех CpG динуклеотидов и в большинстве случаев не метилированы. Если CpGi находится на территории промотора гена, то его метилирование приводит к ингибированию функции этого гена. Если же CpGi находится в другой части гена, например, в районе энхансера (англ. enhancer — усилитель) — небольшого участка ДНК, способного связываться с факторами транскрипции и таким образом увеличивать уровень транскрипции генов — то это метилирование может стимулировать транскрипцию.([32] [33]) Что такое ПЦР? Суть метода Полимера́зная цепна́я реа́кция (ПЦР) — экспериментальный метод молекулярной биологии, позволяющий добиться значительного увеличения малых концентраций определённых фрагментов нуклеиновой кислоты (ДНК) в биологическом материале (пробе).
Помимо амплификации ДНК, ПЦР позволяет производить множество других манипуляций с нуклеиновыми кислотами (введение мутаций, сращивание фрагментов ДНК) и широко используется в биологической и медицинской практике, например, для диагностики заболеваний (наследственных, инфекционных), для установления отцовства, для клонирования генов, выделения новых генов. Метод основан на многократном избирательном копировании определённого участка нуклеиновой кислоты ДНК при помощи ферментов в искусственных условиях (in vitro). При этом происходит копирование только того участка, который удовлетворяет заданным условиям, и только в том случае, если он присутствует в исследуемом образце. В отличие от амплификации ДНК в живых организмах, (репликации), с помощью ПЦР амплифицируются относительно короткие участки ДНК. В обычном ПЦР-процессе длина копируемых ДНК-участков составляет не более 3000 пар оснований (3 kbp[8]). С помощью смеси различных полимераз, с использованием добавок и при определённых условиях длина ПЦР-фрагмента может достигать 20—40 тысяч пар нуклеотидов. Это всё равно значительно меньше длины хромосомной ДНК эукариотической клетки. Например, геном человека состоит примерно из 3 млрд пар оснований[9]. ПЦР проводят в амплификаторе — приборе, обеспечивающем периодическое охлаждение и нагревание пробирок, обычно с точностью не менее 0,1 °C. Современные амплификаторы позволяют задавать сложные программы, в том числе с возможностью «горячего старта», Touchdown ПЦР (см. ниже) и последующего хранения амплифицированных молекул при 4 °C. Для ПЦР в реальном времени выпускают приборы, оборудованные флуоресцентным детектором. Существуют также приборы с автоматической крышкой и отделением для микропланшет, что позволяет встраивать их в автоматизированные системы. 27. Алгоритм проведения ПЦР Обычно при проведении ПЦР выполняется 20—35 циклов, каждый из которых состоит из трёх стадий Денатурация Двухцепочечную ДНК-матрицу нагревают до 94—96 °C (или до 98 °C, если используется особенно термостабильная полимераза) на 0,5—2 мин, чтобы цепи ДНК разошлись. Эта стадия называется плавлением (денатурацией), так как разрушаются водородные связи между двумя цепями ДНК. Обычно перед первым циклом проводят длительный прогрев реакционной смеси в течение 2—5 мин для полной денатурации матрицы и праймеров. Отжиг Когда цепи разошлись, температуру понижают, чтобы праймеры могли связаться с одноцепочечной матрицей. Эта стадия называется отжигом. Температура отжига зависит от состава праймеров и обычно выбирается на 5 градусов меньше, чем температура плавления праймеров. Неправильный выбор температуры отжига приводит либо к плохому связыванию праймеров с матрицей (при завышенной температуре), либо к связыванию в неверном месте и появлению неспецифических продуктов (при заниженной температуре). Время стадии отжига — 30 сек, одновременно, за это время полимераза уже успевает синтезировать несколько сотен нуклеотидов. Поэтому рекомендуется подбирать праймеры с температурой плавления выше 60 °C и проводить отжиг и элонгацию одновременно, при 60—72 °C. Элонгация ДНК-полимераза реплицирует матричную цепь, используя праймер в качестве затравки. Это — стадия элонгации. Полимераза начинает синтез второй цепи от 3'-конца праймера, который связался с матрицей, и движется вдоль матрицы, синтезируя новую цепь в направлении от 5' к 3' концу. Температура элонгации зависит от полимеразы. Часто используемые полимеразы Taq и Pfu наиболее активны при 72 °C. Время элонгации зависит как от типа ДНК-полимеразы, так и от длины амплифицируемого фрагмента. Обычно время элонгации принимают равным одной минуте на каждую тысячу пар оснований. После окончания всех циклов часто проводят дополнительную стадию финальной элонгации, чтобы достроить все одноцепочечные фрагменты. Эта стадия длится 7—10 мин. Количество специфического продукта реакции (ограниченного праймерами) теоретически возрастает пропорционально 2n — 2n, где n — число циклов реакции[15]. На самом деле эффективность каждого цикла может быть меньше 100 %, поэтому в действительности P ~ (1+E)n, где P — количество продукта, Е — средняя эффективность цикла. Число «длинных» копий ДНК тоже растет, но линейно, поэтому в продуктах реакции доминирует специфический фрагмент. Рост требуемого продукта в геометрической прогрессии ограничен количеством реагентов, присутствием ингибиторов, образованием побочных продуктов. На последних циклах реакции рост замедляется, это называют «эффектом плато». Компоненты ПЦР Компоненты реакции[править | править вики-текст] Для проведения ПЦР в простейшем случае требуются следующие компоненты: ДНК-матрица, содержащая тот участок ДНК, который требуется амплифицировать. Два праймера, комплементарные противоположным концам разных цепей требуемого фрагмента ДНК. Термостабильная ДНК-полимераза — фермент, который катализирует реакцию полимеризации ДНК. Полимераза для использования в ПЦР должна сохранять активность при высокой температуре длительное время, поэтому используют ферменты, выделенные из термофилов — Thermus aquaticus (Taq-полимераза), Pyrococcus furiosus (Pfu-полимераза), Pyrococcus woesei (Pwo-полимераза), Thermus thermophilus (Tth-полимераза) и другие. Дезоксирибонуклеозидтрифосфаты (dATP, dGTP, dCTP, dTTP). Ионы Mg2+, необходимые для работы полимеразы. Буферный раствор, обеспечивающий необходимые условия реакции — рН, ионную силу раствора. Содержит соли, бычий сывороточный альбумин. Чтобы избежать испарения реакционной смеси, в пробирку добавляют высококипящее масло, например, вазелиновое. Если используется амплификатор с подогревающейся крышкой, этого делать не требуется. Добавление пирофосфатазы может увеличить выход ПЦР-реакции. Этот фермент катализирует гидролиз пирофосфата, побочного продукта присоединения нуклеотидтрифосфатов к растущей цепи ДНК, до ортофосфата. Пирофосфат может ингибировать ПЦР-реакцию[10]. Праймеры Специфичность ПЦР основана на образовании комплементарных комплексов между матрицей и праймерами, короткими синтетическими олигонуклеотидами длиной 18—30 оснований. Каждый из праймеров комплементарен одной из цепей двуцепочечной матрицы и ограничивает начало и конец амплифицируемого участка. Tm — температура, при которой половина ДНК-матриц образует комплекс с олигонуклеотидным праймером. В случае неверного выбора длины и нуклеотидного состава праймера или температуры отжига возможно образование частично комплементарных комплексов с другими участками матричной ДНК, что может привести к появлению неспецифических продуктов. Верхний предел температуры плавления ограничен оптимумом температуры действия полимеразы, активность которой падает при температурах выше 80 °C. ПЦР проводят в амплификаторе — приборе, обеспечивающем периодическое охлаждение и нагревание пробирок, обычно с точностью не менее 0,1 °C. Современные амплификаторы позволяют задавать сложные программы, в том числе с возможностью «горячего старта», Touchdown ПЦР (см. ниже) и последующего хранения амплифицированных молекул при 4 °C. Для ПЦР в реальном времени выпускают приборы, оборудованные флуоресцентным детектором. Существуют также приборы с автоматической крышкой и отделением для микропланшет, что позволяет встраивать их в автоматизированные системы. Ошибки при проведении ПЦР Очень важным моментом является воспроизводимость результатов ПЦР. Для того чтобы этот показатель был высоким, нужно стандартизовать условия проведения ПЦР. Условия должны быть одинаковыми для всех проб. Рассмотрим типичные источники ошибок, встречающиеся на каждом из этапов ПЦР. Этап выделения ДНК. 1. Неправильное хранение биологического материала 2. Загрязнение проб 3. Нарушениие очередности действий в этапе выделения ДНК 4. Истечение срока годности реактивов для выдления ДНК Этапы амплификации ДНК 1. Нарушение пропорций при смешивании реактивов для амплификации 2. Использование праймера, не способного отжигаться на ДНК данного организма 3. Загрязнение проб ДНК из оседних пробирок 4. Истечение срока годности реактивов для амплиикации ДНК Этапы детекции продуктов амплификации 1. Использование силы тока и напряжения, неподходящих для электрофореза 2. Неправильное подключение камеры для электрофореза 3. Испорченный трис-боратный буфер 4. Неправильное приготовление и старение агарозного геля Стадии постановки ПЦР ПЦР-анализ состоит из трех стадий
Подготовка пробы биологического материала. Для выделения ДНК используют различные методики в зависимости от поставленных задач. Их суть заключается в экстракции (извлечении) ДНК из биопрепарата и удалении или нейтрализации посторонних примесей для получения препарата ДНК с чистотой, пригодной для постановки ПЦР. Иногда бывает достаточно прокипятить образец в течение 5-10 мин., однако в большинстве случаев требуются более сложные методы. Стандартной и ставшей уже классической считается методика получения чистого препарата ДНК, описанная Мармуром. Она включает в себя ферментативный протеолиз с последующей депротеинизацией и переосаждением ДНК спиртом. Этот метод позволяет получить чистый препарат ДНК. Однако он довольно трудоемок и предполагает работу с такими агрессивными и имеющими резкий запах веществами, как фенол и хлороформ. Амплификация Для проведения реакции амплификации необходимо приготовить реакционную смесь и внести в нее анализируемый образец ДНК. При этом важно учитывать некоторые особенности отжига праймеров. Дело в том, что, как правило, в анализируемом биологическом образце присутствуют разнообразные молекулы ДНК, к которым используемые в реакции праймеры имеют частичную, а в некоторых случаях значительную, гомологию. Оценка результатов реакции.
Для правильной оценки результатов ПЦР важно понимать, что данный метод не является количественным. Теоретически продукты амплификации единичных молекул ДНК-мишени могут быть обнаружены с помощью электрофореза уже после 30-35 циклов. Однако на практике это выполняется лишь в случаях, когда реакция проходит в условиях, близких к идеальным, что в жизни встречается не часто. Особенно большое влияние на эффективность амплификации оказывает степень чистоты препарата ДНК, т.е. наличие в реакционной смеси тех или иных ингибиторов, от которых избавиться в некоторых случаях бывает крайне сложно. Иногда, из-за их присутствия не удается амплифицировать даже десятки тысяч молекул ДНК-мишени. Таким образом, прямая связь между исходным количеством ДНК-мишени и конечным количеством продуктов амплификации часто отсутствует.
Метод горизонтального электрофореза
Для визуализации результатов амплификации используют различные методы. Наиболее распространенным на сегодняшний день является метод электрофореза, основанный на разделении молекул ДНК по размеру. Для этого готовят пластину агарозного геля, представляющего собой застывшую после расплавления в электрофорезном буфере агарозу в концентрации 1,5-2,5% с добавлением специального красителя ДНК, например, бромистого этидия. Застывшая агароза образует пространственную решетку. При заливке с помощью гребенок в геле формируют специальные лунки, в которые в дальнейшем вносят продукты амплификации. Пластину геля помещают в аппарат для горизонтального гель-электрофореза и подключают источник постоянного напряжения. Отрицательно заряженная ДНК начинает двигаться в геле от минуса к плюсу. При этом более короткие молекулы ДНК движутся быстрее, чем длинные. На скорость движения ДНК в геле влияет концентрация агарозы, напряженность электрического поля, температура, состав электрофорезного буфера и, в меньшей степени, ГЦ-состав ДНК. Все молекулы одного размера движутся с одинаковой скоростью. Краситель встраивается (интеркалирует) плоскостными группами в молекулы ДНК. После окончания электрофореза, продолжающегося от 10 мин до 1 часа, гель помещают на фильтр трансиллюминатора, излучающего свет в ультрафиолетовом диапазоне (254 - 310 нм). Энергия ультрафиолета, поглощаемая ДНК в области 260 нм, передается на краситель, заставляя его флуоресцировать в оранжево-красной области видимого спектра (590 нм). Яркость полос продуктов амплификации может быть различной. Поэтому часто в ПЦР-лабораториях принято оценивать результат по трех- четырех или пяти- балльной системе. Однако, как уже отмечалось ранее, это нельзя связывать с начальным количеством ДНК-мишени в образце. Часто уменьшение яркости свечения полос связано со снижением эффективности амплификации под влиянием ингибиторов или других факторов.
Метод вертикального электрофореза
Метод вертикального электрофореза принципиально схож с горизонтальным электрофорезом. Их отличие заключается в том, что в данном случае вместо агарозы используют полиакриламидные гели. Его проводят в специальной камере для вертикального электрофореза. Электрофорез в полиакриламидном геле имеет большую разрешающую способность по сравнению с агарозным электрофорезом и позволяет различать молекулы ДНК разных размеров с точностью до одного нуклеотида. Приготовление полиакриламидного геля несколько сложнее агарозного. Кроме того акриламид является токсичным веществом. Поскольку необходимость определить размер продукта амплификации с точностью до 1 нуклеотида возникает редко, то в рутинной работе используют метод горизонтального электрофореза.
Метод гибридизационных зондов
Другой способ детекции продуктов амплификации основан на гибридизации олигонуклеотидных зондов с продуктами амплификации, содержащими ту или иную метку. В этом случае чаще всего используют плашечный формат и систему детекции, аналогичную используемой в иммуноферментном анализе (ИФА). Гибридизационные методы не получили пока широкого распространения из-за их дороговизны, длительности и трудоемкости методических манипуляций. Однако они интересны с точки зрения массового скрининга, т.к. при наличии соответствующего оборудования могут быть легко автоматизированы. Вставка гена в вектор Ген вводится в специальные молекулы ДНК, которые обеспечивают функционирование гена в новой генно-инженерной системе. Генетические элементы, используемые для создания рекомбинантной ДНК и введения ее в клетку, называют векторами. В качестве векторов чаще всего используют вирусы и плазмиды. Плазмидные векторы созданы на основе природных бактериальных плазмид. Они представляют собой небольшие кольцевые молекулы двухцепочечной ДНК. Такие векторы способны автономно реплицироваться благодаря наличию точки инициации репликации (ori). Плазмиды, которые используются в генной инженерии, содержат селективные маркеры. В качестве селективных маркеров часто используются гены устойчивости к антибиотикам. Обычно векторная молекула несет более одного селективного маркера. Благодаря таким маркерным генам на последующих этапах клетки, несущие рекомбинантную плазмиду, легко отделить от клеток, не имеющих такой рекДНК. Вставка гена в вектор, т.е. создание рекДНК осуществляется стандартно с использованием рестриктаз. Векторная молекула, кроме oriи селективных маркеров должна нести уникальные сайты рестрикции для нескольких рестриктаз. Уникальный сайт рестрикции обеспечивает то, что при обработке соответствующей рестриктазой кольцевая молекула будет разрезана в одном строго определенном месте и превратится в линейную. При соединении клонируемого гена и вектора, обработанных одной и той же рестриктазой, эти молекулы могут соединяться в одно целое путем спаривания оснований «липких концов». Полученная молекула ДНК (вектор со встроенным геном) называется рекомбинантной (рекДНК). Соответственно, кодируемые рекомбинантными ДНК ферменты называютрекомбинантными ферментами, а клетки, экспрессирующие такие ферменты (или другие белки) – рекомбинантными клетками или рекомбинантными системами. Отбор клонов Все клетки одного клона генетически однородны и могут быть нескольких вариантов: · не содержать рекДНК, · содержать вектор без вставки гена; · содержать рекДНК. Для того, чтобы выявить клоны, содержащие рекДНК используется отбор на основе селективных маркеров. Так, когда в качестве селективных маркеров используется гены устойчивости к антибиотикам, клетки высевают на селективную питательную среду, содержащую соответствующий антибиотик. В таком случае расти будут только те клетки, которые имеют вектор с геном устойчивости. Чтобы выявить клетки, несущие не просто вектор, а рекДНК, используется отбор на основании второго маркера. В случае клонирования фрагментов, полученных из геномной ДНК или кДНК получают набор клонов, несущих рекДНК с различными вставками – библиотеки геномной и кДНК. Для отбора клона с нужным геном из библиотеки используется гибридицация нуклеиновых кислот или анализ с помощью антител к белковому продукту клонированного гена. Функционирование гена в клетке-реципиенте (адаптация гена) Плазмида, попадающая в клетку-хозяин и придающая ей новые свойства является нестабильным образованием. Возможна ее деструкция под действием нуклеаз. Кроме того, на ее стабильность оказывает влияние динамика роста всей клеточной популяции. Выявляются определенные кинетические закономерности репликации плазмид. Число плазмид на клетку не является постоянным, а закономерно изменяется в процессе роста популяции. При этом все наблюдаемые случаи могут быть разделены на два типа – монофазные и двухфазные процессы (рис.5.1.1). В первом случае клетки временно теряют некоторое количество плазмид. Второй случай более сложен: уменьшение числа плазмид на начальном этапе в последующем сопровождается их гиперпродукцией. Считается, что особенности кинетики репликации плазмид первого и второго типа обусловлены наличием периода индукции в случае плазмид второго типа. Клонирование ДНК Клонирование - один из методов выделения и идентификации фрагментов ДНК, а также получения их в неограниченном количестве. Выделенные и идентифицированные фрагменты могут быть использованы для молекулярного анализа, в качестве ДНК-зондов и ддя создания библиотек генов. Метод был разработан на бактериях и свое название получил из-за того, что все производные одной бактериальной колонии содержат идентичные фрагменты ДНК, т.е. представляют собой клоны. Клонирование фрагмента ДНК включает несколько последовательных этапов: 1) встраивание клонируемого (чужеродного) фрагмента ДНК в векторную молекулу ДНК (образование химерной молекулы - рекомбинантной ДНК); 2) проникновение этой конструкции в бактериальную клетку-хозяина; 3) идентификация клеток, содержащих рекомбинантную ДНК, и их отбор (как правило, осуществляется на селективной среде); 4) получение необходимого количества клеток, содержащих рекомбинантную ДНК (собственно клонирование). При необходимости индуцируют экспрессию клонированного гена в клетках-хозяевах и получают кодируемый им белок. Итак, клонирование предполагает встраивание (инсерцию) экзогенной ДНК в векторную молекулу ДНК. Векторные системы, обеспечивающие доставку чужеродного фрагмента ДНК в клетку хозяина, для прокариот и эукариот различны. Для клонирования в прокариотических клетках используют плазмиды, фаги и космиды. В зависимости от типа векторной системы, используемой для доставки клонируемого фрагмента ДНК, процесс переноса генов носит название: при использовании плазмиды — трансформации; при использовании фага - трансдукции. Перенос экзогенной ДНК в эукариотические клетки называют трансфертен. В качестве векторов для переноса ДНК в эукариотические клетки используют дрожжевые плазмиды (единственные плазмиды, найденные в эукариотических клетках) и различные эукариотические вирусы (чаще всего ретровирусы, аденовирусы или аденоассоциированные вирусы). В ряде случаев введение векторных конструкций в эукариотические клетки осуществляют путем ко-трансформацый — одновременного введения плазмиды и сегмента чужеродной ДН К. В клетках эукариот векторные конструкции сохраняются в виде эписом (суперскрученных кольцевых молекул) в течение нескольких дней, а иногда экзогенная ДНК интегрируется в хромосомную ДНК и устойчиво сохраняется в геноме клетки-хозяина. Геномные библиотеки Рестрикция геномной ДНК на фрагменты и клонирование фрагментов с помощью различных векторов создали основу формирования геномных библиотек. Для этого геномная ДНК разрезается или, как говорят, переваривается определенной рестриктазой, а образующиеся фрагменты клонируются с помощью различных векторов, для чего используют методы рекомбинантной ДНК. Геномная библиотека должна содержать не только гены, но и всю некодирующую ДНК, расположенную между генами. Поскольку переваривание рестриктазой производят неполное так, что одни сайты, специфические для рестриктазы, разрезаются, а другие нет, то образуются фрагменты ДНК с частично перекрывающимися последовательностями нуклеотидов. Это облегчает последующее восстановление картины расположения фрагментов в нативной ДНК. Кроме геномных библиотек, существуют библиотеки кДНК. Библиотеки кДНК содержат только экзоны генов, поскольку получаются на основе зрелой мРНК с помощью фермента, называемого обратной транскриптазой. Обратная транскриптаза на матрице мРНК создает комплементарную нить ДНК, которая затем превращается в обычную двунитевую ДНК с помощью ДНК-полимеразы. Затем такие молекулы кДНК клонируются в бактериальных клетках точно так же, как и геномная ДНК. Еще один тип ДНК-библиотек — хромосомоспецифические библиотеки. Для их создания хромосомы разделяют с помощью проточной цитометрии, которая позволяет выделить отдельные хромосомы. ДНК, полученная после такой сортировки, будет преимущественно представлять определенную хромосому. Затем получают фрагменты ДНК отдельной хромосомы перевариванием с той или иной рестриктазой и клонируют их обычным путем. Различные библиотеки ДНК широко используют при реализации программы «Геном человека», а также для других целей, например при поиске полиморфных ДНК-маркеров. Этапы клонирования ДНК Клонирование фрагмента ДНК включает несколько последовательных этапов: 1) встраивание клонируемого (чужеродного) фрагмента ДНК в векторную молекулу ДНК (образование химерной молекулы - рекомбинантной ДНК); 2) проникновение этой конструкции в бактериальную клетку-хозяина; 3) идентификация клеток, содержащих рекомбинантную ДНК, и их отбор (как правило, осуществляется на селективной среде); 4) получение необходимого количества клеток, содержащих рекомбинантную ДНК (собственно клонирование). При необходимости индуцируют экспрессию клонированного гена в клетках-хозяевах и получают кодируемый им белок. Векторы клонирования Типы векторов для введения гена в клетку
Существует несколько типов векторов: Бактериальные плазмиды Основная масса клеточной ДНК бактерий содержится в хромосоме (в хромосоме E. coli, например, 4 млн. пар нуклеотидов). Однако кроме хромосом бактерии содержат большое количество очень маленьких кольцевых молекул ДНК плазмид длиной несколько тысяч пар оснований (молекулярная масса от 1,5 до 300 мегадальтон, 1 МД = 1500 п.о). Такие мини-хромосомы называют плазмидами. Как правило, плазмиды имеют в своем составе гены устойчивости к антибиотикам, ионам тяжелых металлов (R-плазмиды), а также гены, контролирующие катаболизм некоторых органических соединений (плазмиды биодеградации, или D-плазмиды). Поскольку эти гены находятся в плазмидах, они представлены гораздо большим числом копий. Высокая копийность плазмид обеспечивает клетке синтез большого количества ферментов, химически нейтрализующих антибиотики или ксенобиотики, что и обеспечивает устойчивость к последним. Плазмиды, по-видимому, вездесущи, так как их выделяют из разных штаммов и видов бактерий, но не являются обязательными компонентами генома, а в некоторых природных штаммах плазмиды не обнаружены вообще. Поскольку плазмидная ДНК значительно меньше хромосомной, ее довольно легко выделить в чистом виде. В присутствии ионов кальция плазмиды легко поглощаются бактериями-рецепиентами, даже если те их никогда не содержали, и в клетках бактериального потомства можно обнаружить много копий поглощенной плазмиды. Однако бактериальная клетка обычно может содержать в своем составе плазмиды одного типа. Это явление несовместимости плазмид. Существуют группы несовместимости – Inc-группы (от английского incompatibility – несовместимость). В такой группе может быть несколько плазмид, совместимых между собой, но не совместимых с другими плазмидами. У этих плазмид сходны многие признаки и часто значительна гомология ДНК. Число копий плазмиды в клетке может существенно варьировать. Это зависит от генетических особенностей как клетки, так и плазмиды. Плазмиды, находящиеся "под ослабленным контролем", могут размножаться до тех пор, пока их количество не достигнет 10-200 копий на клетку. Если же плазмида находится "под строгим контролем", она реплицируется с той же скоростью, что и главная хромосома. Такие плазмиды содержатся в клетке в одной или в нескольких копиях. Естественно, что для клонирования рекомбинантных ДНК стараются использовать плазмиды первого типа. Но это не обязательно, так как плазмиды в присутствии хлорамфеникола могут умножаться независимо от деления хромосомы, и количество копий плазмиды может многократно увеличиваться. Одна их наиболее часто употребляемых плазмид для клонирования pBR 322 создана на основе плазмид природного происхождения, выделенных из E. coli. Эта плазмида содержит гены устойчивости к двум антибиотикам: ампициллину и тетрациклину, причем в генах устойчивости к этим антибиотикам имеются сайты рестрикции. Если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику. Но при этом сохраняется устойчивость к другому антибиотику. Таким образом вектор дает возможность детектировать только те клоны бактерий, которые содержат рекомбинантную плазмиду. Вирусы Есть вирусы, которые не ведут к гибели клетки, но встраиваются в геном клетки-хозяина и размножаются вместе с ней, либо вызывают ее неконтролируемый рост, т.е. превращают в раковую. К таким относятся ДНК-вирусы SV-40 и вирус полиомы. Внедрение некоторых опухолевых РНК-вирусов ведет к отпочковыванию вирусных частиц от клетки без ее лизиса. К таким вирусам относятся, например, ретровирусы (вирус саркомы Рауса и СПИДа). Для бактериальных клеток в качестве вектора часто используют бактериофаги. Вирусы являются одними из главных кандидатов на роль векторов для введения чужеродной ДНК. При вирусной инфекции каждая клетка может получить большое число копий чужеродного гена. ДНК можно встраивать так, чтобы она находилась под контролем сильных вирусных промоторов, что обеспечит высокий уровень экспрессии гена, и его продукты будут более доступны для исследования. В последние годы сконструированы многочисленные "челночные" векторы и их рекомбинантные производные, способные к репликации в животной и бактериальной клетке и эффективно экспрессирующие клонируемый ген в животной клетке. Наиболее распространенные векторы состоят из плазмиды рВR322 и интактного раннего района транскрипции ДНК SV40, а нужный ген встраивается под контроль промотора поздних генов или дополнительного раннего промотора. Например, в ДНК SV40 был встроен ген β-глобина кролика, который экспрессировался в линии клеток обезьяны, зараженных рекомбинантным вирусом: в клетках синтезировались и мРНК гена глобина, и сам белок. Вирус должен быть жизнеспособным после рекомбинирования его ДНК. Легче всего вирусы вводятся в бактерии. Недостатком вирусов как векторов является их небольшая емкость. Кроме того, вирусы заражают небольшой круг хозяев. Существуют гибридные вектора, содержащие ДНК фага и плазмиды. К ним относятся, например, космиды и фазмиды. Космиды – плазмидные вектора, в которые встроен участок генома фага λ, обеспечивающий возможность упаковки этой молекулы ДНК в фаговую частицу. Фаговые частицы обеспечивают хорошее проникновение гибридной ДНК в клетку (путем инъекции), после чего происходит замыкание ДНК в кольцо по липким концам и репликация ее по плазмидному типу. Фазмиды также являются гибридами между фагом и плазмидой. После встройки чужеродной ДНК могут в одних условиях развиваться как фаги, в других – как плазмиды. Вироиды Из всех известных в настоящее время инфекционных агентов имеют ранг наиболее странных. Известно, что самые мелкие вирусы, способные к независимой репликации, имеют размеры генома, соответствующие молекулярной массе 1 М, то есть около 1500 тыс. пар оснований. Это считали минимальным количеством генетической информации, необходимой для кодирования вирусоспецифических продуктов и подавления метаболизма хозяйской клетки. Однако в 1971 году были открыты инфекционные агенты, представляют собой очень короткую цепь 1 нитевой ковалентно связанной кольцевой РНК, состоящую из 270-300 нуклеотидов (на три порядка меньше самых минимальных вирусов), не заключенную в белковую оболочку. Это необычные патогены - самые простые и самые маленькие из всех известных. Каким образом вироиды продуцируют симптомы болезни в инфицированных растениях, не известно до сих пор. Установлено, что они реплицируются ферментами клетки-хозяина, не транслируются в видоспецифичные полипептиды, интегрируются в геном клетки-хозяина. Вироиды заражают персиситентно (не происходит выздоровления). Вызывают системную инфекцию, т.е. мигрируют из сайта внедрения в другие части растений, переносятся механически или через клеточный сок, через семена, пыльцу. Вироиды также связаны с ядерными фракциями растений и могут размножаться в ядрах. При работе с вироидами получают 1-нитевую ДНК- копию РНК и достраивают комплементарную нить для получения 2-нитевой ДНК вироида. Такая 2-цепочечная ДНК вcтраивается в плазмиду и передается в клетки E. coli для клонирования. Считывание гена начинается с промотора, который узнается РНК-полимеразой, отвечающей за транскрипцию ДНК в матрицу РНК. Обычно это фрагмент ДНК из 41-44 пар оснований. Ген считывается слева направо, от 5’ к 3’ концу гена и заканчивается в терминальной области гена. За промотором начинается стартовый сайт транскрипции, за которым следует смысловая часть гена. Промоторная область гена содержит определенные короткие сочетания нуклеотидов, характерные для бактериальных генов, или для генов высших организмов. Такие сочетания служат сигналами для РНК-полимеразы, которая присоединяется к промоторной части гена и начинает его считывать. Однонитевые и двунитевые ДНК способны инициировать репликацию вироида в механически инокулированных растениях табака. Энзиматически in vitro синтезированы также РНК вироидов, высокоинфекционные для растений. Векторные системы могут быть разработаны на основе самих РНК, на основе вироидоспецифичных ДНК, а также в комбинации вироидоспецифичных ДНК с Ti-плазмидами. Вироиды инфицируют своих хозяев в течение всего их жизненного цикла, поэтому в случае использования вироидных векторных систем можно ожидать постоянной экспрессии чужеродного гена в растении. Плазмиды агробактерий В качестве векторов могут использоваться опухолеобразующие плазмиды бактерий. Виды Agrobacterium эволюционно родственны клубеньковым бактериям, относящимся к роду Rhizobium, и имеют много общих с ними черт. Однако характер взаимодействия агробактерий с растением имеет своеобразные особенности. Взаимодействие видов Agrobacterium с растениями представляет особый интерес, так как при этом виде паразитизма один из партнеров специфически видоизменяет свойства хозяина, встраивая свои гены в его геном. Кроме того, это служит уникальным примером миграции ДНК прокариот в эукариотическую клетку. ДНК митохондрий и хлоропластов Хлоропласты и митохондрии содержат полноценную генетическую систему, то есть все компоненты, необходимые для экспрессии генетической информации: ДНК, ДНК-полимеразы, РНК-полимеразы и белоксинтезирующий аппарат (рибосомы, т-РНК, аминоацил-тРНК-синтетазы). Хлоропластная и митохондриальная ДНК также привлекают внимание ученых в качестве возможных векторов для переноса генов в клетку. Структурная организация этих клеточных субгеномов существенно различается. Хлоропласты и другие пластиды обладают одинаковой генетической информацией, так называемым пластомом. У высших растений он представляет собой замкнутую молекулу ДНК длиной 150 т. н. п., достаточную для кодирования примерно 100 белков. Для синтеза пластид необходимо значительно больше белков. Остальные белки кодируются ядром, синтезируются в цитоплазме и поступают в хлоропласты. Некоторые важнейшие белки хлоропластов состоят из нескольких субъединиц, часть из них синтезируется на рибосомах цитоплазмы и транспортируется в хлоропласт, где они объединяются с другими полипептидами, закодированными в самом хлоропласте и там же синтезируемыми. Таким образом, для биосинтеза функционально активного хлоропласта требуется согласованная экспрессия генома и пластома. Различные типы пластид содержат неодинаковые количества идентичных копий пластома: от 10 – 20 копий в пластидах корней и зрелых хлоропластах до сотен копий в молодых хлоропластах картофеля. Такой уровень амплификации позволяет надеяться на надежную экспрессию чужеродной ДНК при использовании их в качестве векторов в генноинженерных экспериментах. Кроме того, гены рибосомальной РНК пластид и большой субъединицы РБФК кодируются геномом хлоропластов. Возможно, введение сильных промоторов в эти гены и дополнительная их модификация существенно повлияют на фотосинтетическую активность растительной ткани. Гены растений также способны к экспрессии в клетках Е. coli. Это гены большой субъединцы РБФК. Преимущество хлоропластных генов заключается в том, что их экспрессия к клетках кишечной палочки может быть достигнута путем простого объединения транскрибируемых последовательностей, т.к. в ДНК хлоропластов и бактерий до начала стартовых кодонов трансляции расположена одинаковая нуклеотидная последовательность. Это дает возможность синтезировать растительные экономически важные полипептиды с помощью клеток прокариот. В отличие от хлоропластной, ДНК митохондрий характеризуются исключительным разнообразием и их величина колеблется от 200 до 2400 т. н. п.. Однако никакой корреляции между размером митохондриального генома и числом белковых продуктов, синтезируемых изолированными митохондриями, не наблюдается. Это явление, а также большие размеры митохондриальной ДНК, по-видимому, можно объяснить присутствием ДНК, бесполезной для функционирования митохондрий. В составе митохондриальной ДНК имеются структурные гены, кодирующие полипептиды, гены рибосомных и транспортных РНК. Однако большая часть белков митохондрий, как и хлоропластов, кодируется ядерными генами. Но если геном хлоропластов представлен гомогенной популяцией крупных кольцевых молекул, то в митохондриях содержится несколько классов кольцевых молекул, не все функции которых еще ясны. Митохондриальный геном животных организмов намного меньше, 15 – 19 т. н. п., и более консервативен по структуре. Гены митохондрий кодируют 2 группы признаков – работу дыхательных систем и устойчивость к антибиотикам и другим ядам. В митохондриальном геноме растений есть также гены, отвечающие за признак мужской стерильности цитоплазмы. Транспозоны Транспозоны - сегменты ДНК, которые контролируют собственную транспозицию (перемещение) из одного сайта ДНК в другой путем вырезания из исходного сайта и внедрения в новый сайт хромосомы или плазмиды. Впервые были открыты в 40-х годах американской ученой Барбарой Мак-Клинток у кукурузы. Эти гены, индентифицированные по их способности подавлять экспрессию других генов кукурузы, находящихся рядом с ними, не имели фиксированного положения в хромосоме. Они как бы передвигались по всему геному растения. Регуляторные элементы могли встраиваться и выщепляться, причем после их выщепления зачастую начинали функционировать ранее молчащие гены. Оказалось, что гены, ассоциированные с регуляторными элементами, становились нестабильными и часто мутировали из-за нестабильности самих этих элементов. В течение многих лет кукуруза оставалась единственной системой, в которой обнаруживались такие подвижные генетические элементы. Сейчас - и у бактерий, дрозофил и других организмов. Механизм перемещения фрагментов ДНК по геному до конца не выяснен. ДНК переносится ферментом транспозазой. Фермент кодируется последовательность длиной около 20 нуклеотидов в середине транспозона. Он специфически взаимодействует с концевыми инвертированными повторами мобильного элемента и может вырезать его из хромосомы. Вырезание может происходить точно – с восстановлением исходной структуры участка ДНК, и неточно, то есть с делециями и вставками от одного до нескольких нуклеотидов. Это приводит к появлению стабильных мутаций и является одним из механизмов создания новых последовательностей ДНК. Как правило, мобильные генетические элементы многократно повторены в геноме и образуют гетерогенные семейства, члены которых диспергированы по хромосомам. Большая часть членов каждого семейства являются дефектными копиями и не кодируют какой-либо функции, хотя сохраняют способность к перемещению. Поведение транспозонов можно расценить как паразитическое. Длина их от 2 до 10 тысяч нуклеотидных пар. У высших эукариот на долю транспозонов приходится примерно 10% ДНК клетки. Большинство их перемещается изредка, но, так как их в клетке довольно много, транспозиция оказывает значительное влияние на разнообразие видов. Биологический смысл перемещения отдельных сегментов ДНК: - прерывание соответствующего гена, что ведет к эволюции; - регуляция деятельности генов, так как транспозоны могут нести сигналы для начала считывания генов. В новых областях усиливают или запрещают работу гена. Транспозоны также участвуют в горизонтальном переносе генов. У бактерий были обнаружены 2 класса подвижных генов, различающихся по длине и сложности организации. 1. Инсерционные последовательности, или 1S элементы, имеющие длину около тысячи пар нуклеотидов и содержащие только ген, отвечающий за их перемещение. 2. Транспозоны, длиной от 3 до 20 т. н. п., состоящие из ряда дополнительных генов, отвечающих за устойчивость бактерий к различным токсическим веществам. Поскольку подвижные гены могут перемещаться в пределах генома с одного места на другое, то они могут быть весьма эффективными векторами для передачи рекомбинантной ДНК. Генетическая трансформация с помощью векторов на основе транспозонов была впервые осуществлена на дрозофиле. С помощью транспозируемого элемента Р дрозофиле был передан ген, обуславливающий коричневую окраску глаз. Перенос генов при помощи транспозонов имеет большие преимущества, так как он происходит с высокой частотой и не влечет значительных перестроек интегрируемой ДНК. Кроме того, этим методом можно переносить достаточно большие фрагменты ДНК. Базы данных НК и белков BLAST (англ. Basic Local Alignment Search Tool - средство поиска основного локального выравнивания) — семейство компьютерных программ, служащих для поиска гомологов белков или нуклеиновых кислот, для которых известна первичная структура (последовательность) или её фрагмент. Используя BLAST, исследователь может сравнить имеющуюся у него последовательность с последовательностями из базы данных и найти последовательности предполагаемых гомологов. После введения изучаемой нуклеотидной или аминокислотной последовательности (запрос) на одну из веб-страниц BLAST, она вместе с другой входной информацией (база данных, размера «слова» (участка), значение величины E и др.) поступает на сервер. BLAST создаёт таблицу всех «слов» (в белке — это участок последовательностей, который по умолчанию состоит из трёх аминокислот, а для нуклеиновых кислот из 11 нуклеотидов) и сходных «слов». Затем в базе данных проводится их поиск. Когда обнаруживается соответствие, то делается попытка продлить размеры «слова» (до 4 и более аминокислот и 12 и более нуклеотидов) сначала без гэпов (пробелов), а затем с их использованием. После максимального продления размеров всех возможных «слов» изучаемой последовательности, определяются выравнивания с максимальным количеством совпадений для каждой пары запрос — последовательность базы данных, и полученная информация фиксируется в структуре SeqAlign. Форматер, расположенный на сервере BLAST, использует информацию из SeqAlign и представляет её различными способами (традиционным, графическим, в виде таблицы). Для каждой обнаруженной в базе данных программами BLAST последовательности необходимо определить, насколько она сходна с изучаемой последовательностью (запрос) и значимо ли это сходство. Protein Data Bank, PDB — банк данных 3-D структур белков и нуклеиновых кислот. Информация, полученная методами рентгеновской кристаллографии или ЯМР-спектроскопии, вносится в базу данных биологами и биохимиками со всего мира, и доступна бесплатно через интернет. Изомеризация пролина Цис-транс изомеризация Пептид связывается с пролином и с другими N-замещенными аминокислотами (такими, как саркозин), и способен заполнять как цис-, так и транс- изомеры. Большинство пептидных связей принимают транс-изомер (в естественных условиях обычно в 99,9% случаев), главным образом потому, что амид водород (транс-изомер) имеет меньшее стерическое отталкивание с предыдущим \ mathrm {C} ^ {\} альфа атомом, чем со следующим \ mathrm {C} ^ {\} альфа атомом (цис-изомер). В противоположность этому, цис-и транс-изомеры X-Pro пептидной связи (где Х представляет собой любую аминокислоту) испытывают стерические столкновения с соседними замещениями и энергетически почти равны. Таким образом, доля X-Pro пептидных связей в цис-изомере в естественных условиях составляет от 10 до 40%, эта доля слабо зависит от предыдущей аминокислоты с ароматическими остатками в пользу цис-изомера. С кинетической точки зрения, цис-транс-изомеризации пролина - очень медленный процесс, который может препятствовать прогрессу сворачивания белка путем захвата одного или более остатков пролина, что имеет решающее значение для складывания ненативных изомеров, особенно когда нативный белок требует наличия цис-изомеров. Это происходит потому, что остатки пролина синтезируются исключительно в рибосоме, как транс-форма изомера. Все организмы имеют ферменты пролил изомеразы, катализирующие эту изомеризацию, а некоторые бактерии имеют специальные пролил изомеразы, связанные с рибосомой. Однако не все пролины необходимы для фолдинга. Фолдинг (складывание) белков может протекать с нормальной скоростью, несмотря на наличие ненативных конформеров на многих X-Pro пептидных связях. Генетическая трансформация Генети́ческая трансформа́ция — изменение наследственных свойств клетки в результате внесения в нее генетической информации при помощи чужеродной изолированной дезоксирибонуклеиновой кислоты (ДНК). Трансформация приводит к появлению у трансформированной клетки (трансформанта) и ее потомства новых признаков, характерных для объекта — источника ДНК. Явление трансформации было открыто в 1928 году британским ученым Ф. Гриффитом, который показал возможность превращения непатогенных штаммов вирусов Streptococcus pneumoniae в патогенные штаммы, в результате взаимодействия с убитыми клетками патогенных штаммов. В 1944 году американец О. Эйвери установил, что трансформацию обеспечивают молекулы ДНК, которые являются носителями наследственной информации. Первоначально трансформация была изучена на примере бактерий пневмококков. Использование в экспериментах легко учитываемых генетических признаков (например, устойчивость к действию клеточных ядов, потребность в определенных факторах роста), а также применение ДНК с радиоизотопной меткой позволили дать трансформации количественную оценку. Трансформацию у бактерий рассматривают как сложный процесс, включающий стадии: фиксация молекул ДНК клеткой-реципиентом; проникновение ДНК внутрь клетки; включение фрагментов трансформирующей ДНК в хромосому клетки-хозяина; формирование «чистых» трансформированных вариантов. Фиксация ДНК происходит на особых участках клеточной поверхности (рецепторах), число которых ограничено. Связанная с рецепторами ДНК сохраняет чувствительность к действию добавленного в среду фермента дезоксирибонуклеазы, вызывающего ее распад. Спустя короткий срок (в пределах одной минуты) после фиксации, часть ДНК проникает в клетку. Бактериальные клетки одного и того же штамма различаются по проницаемости для ДНК. Клетки бактериальной популяции, способные включать чужеродную ДНК, называются компетентными. Число компетентных клеток в популяции незначительно и зависит от генетических особенностей бактерий и фазы роста бактериальной культуры. Развитие компетенции связывают с синтезом особого белка, обеспечивающего проникновение ДНК в клетку. Поскольку в компетентную клетку может одновременно проникнуть ряд фрагментов трансформирующей ДНК, суммарная величина поглощенной ДНК может быть примерно равна размерам хромосомы клетки-хозяина. После проникновения в клетку двунитевой ДНК одна нить распадается до мононуклеотидов и олигонуклеотидов, вторая — встраивается в хромосому клетки-хозяина путем ее разрывов и воссоединений. Последующая репликация такой гибридной структуры приводит к выщеплению «чистых» клонов трансформантов, в потомстве которых закреплен признак, кодируемый включившейся ДНК. Применение трансформации позволило провести генетический анализ бактерий, у которых не описано иных форм генетического обмена (конъюгации, трансдукции). Разработка метода трансформации у кишечной палочки позволила использовать для трансформации не только фрагменты бактериальной хромосомы, но и ДНК бактериальных плазмид и бактериофагов. Этот метод используется для внесения в клетку гибридной ДНК в исследованиях по генной инженерии. В 1960-х годах началось изучение трансформации у животных, в конце 1970-х годов — у растений. Сравнение Саузерн блоттинг
Саузерн-блоттинг (от англ. Southern blot) — метод, применяемый в молекулярной биологии для выявления определенной последовательности ДНК в образце. Метод Саузерн-блоттинга сочетает электрофорез в агарозном геле для фракционирования ДНК с методами переноса разделённой по длине ДНК на мембранный фильтр для гибридизации. Метод называется по имени изобретателя, английского биолога Эдвина Саузерна.[1]
Другие технологии переноса (блота), например, вестерн-блоттинг, нозерн-блоттинг, саузвестерн-блоттинг используют сходные методы, но для определения РНК или белка в образце, и называются по образцу метода, придуманного Саузерном. Так как Саузерн-блот назван по имени ученого, термин пишут с заглавной буквы, в то время как вестерн-блот и нозерн-блот — со строчной буквы. 1. Рестрикция эндонуклеазами рестрикции для разрезания высокомолекулярной ДНК на более мелкие фрагменты. 2. Фрагменты ДНК подвергаются электрофорезу в агарозном геле для разделения по длине. 3. В случае, если некоторые фрагменты ДНК длиннее 15 кб, перед переносом гель обрабатывают, например, соляной кислотой, которая вызывает депуринизацию ДНК и облегчает перенос на мембрану. 4. В случае, когда используют щелочной метод переноса, агарозный гель помещают в щелочной раствор, при этом двойная спираль ДНК денатурирует и облегчает связывание отрицательно заряженной ДНК с положительно заряженной мембраной для дальнейшей гибридизации. При этом разрушаются и остатки РНК. 5. Листок нитроцеллюлозной (или нейлоновой) мембраны помещают сверху или снизу от агарозного геля. Давление осуществляют непосредственно на гель или через несколько слоев бумаги. Для успешного переноса необходим плотный контакт геля и мембраны. Буфер переносится капиллярными силами из участка с высоким содержанием воды в зону с низким содержанием воды (мембрана). При этом осуществляется перенос ДНК из геля на мембрану. Полианионная ДНК связывается с положительно заряженной мембраной силами ионообменных взаимодействий. 6. Для окончательного закрепления ДНК на мембране, последняя нагревается в вакууме до температуры 80 °C в течение двух часов или освещается ультрафиолетовым излучением (в случае нейлоновых мембран). 7. Осуществляют гибридизацию радиоактивно (флюоресцентно) меченной пробы с известной последовательностью ДНК с мембраной. 8. После гибридизации избыток пробы отмывают с мембраны и визуализируют продукты гибридизации путём авторадиографии (в случае радиоактивной пробы) или оценивают окраску мембраны (в случае использования хромогенного окрашивания).
Норзерн блоттинг Нозерн-блот (англ. Northern blot) — метод исследования экспрессии генов путём тестирования молекул РНК (мРНК) и их фрагментов в образцах.
Метод Нозерн-блот был предложен в 1977 году сотрудниками Стэнфордского университета Джеймсом Олвайном, Дэвидом Кемпом и Джорджем Старком[1] и назван по его аналогии с Саузерн-блот — первым из методов этого типа, предложенным Эдвином Саузерном[2]. Основным отличием метода нозерн-блот от Саузерн-блот является то, что определяемым субстратом является не ДНК, а РНК. Это обуславливает различия в методике — вместо нитроцеллюлозного используется фильтр из диазобензилоксиметил-целлюлозы, в качестве зондов используют комплементарные молекулы ДНК и т. д. Вестерн блоттинг Вестерн-блоттинг (вестерн-блот, белковый иммуноблот, англ. Western blot) — аналитический метод, используемый для определения специфичных белков в образце. На первом этапе использует электрофорез белков в полиакриламидном геле для разделения денатурированных полипептидов по длине (как правило, в присутствии SDS) или по трехмерной структуре белка (в нативном состоянии). Далее белки переносят на нитроцеллюлозную или PVDF мембрану, затем детектируют с использованием антител, специфичных к заданному белку.[1][2]
Существует множество коммерческих компаний, специализирующихся на производстве антител (как моноклональных так и поликлональных) к десяткам тысяч различных белков.[3]
Вестерн-блоттинг используется в молекулярной биологии, биохимии, генетике и в других естественно-научных дисциплинах. В методе Вестерн-блоттинг (Western blotting) на первом этапе смесь белков разделяется методом электрофореза в присутствии додецилсульфата натрия (ДСН), затем переносится на нитроцеллюлозную мембрану методом электроблоттинга. Суть данного метода заключается в том, что гель после электрофореза помещается на нитроцеллюлозную мембрану между слоями фильтровальной бумаги. Собранный таким образом «сэндвич» помещается в электрическое поле так, что комплексы белок-ДСН движутся поперек пластины геля и иммобилизуются (в результате неспецифической сорбции) на поверхности нитроцеллюлозной мембраны. В связывании комплекса белок-ДСН с нитроцеллюлозной мембраной принимают участие в основном силы электрической природы, причем данное взаимодействие является многоточечным и приводит к «распластыванию» белков на поверхности мембраны. Таким образом, после электропереноса мы получаем на нитроцеллюлозе реплику геля с белками, расположенными так же, как и в полиакриламидном геле. После проведения ДСН — электрофореза, электропереноса и сорбции белков из геля на нитроцеллюлозную мембрану третичная конформация белка сильно изменена, если вообще правильно говорить о существовании третичной структуры для белка после такой жесткой обработки. Поэтому для иммунохимической детекции исследуемого белка обычно используются только моно- или поликлональные антитела, специфичные к линейным участкам белковой молекулы. Антитела, специфичные к конформационным эпитопам (или участкам, включающим в себя межсубъединичные контакты), как правило, не пригодны для использования в методе Вестерн-блоттинг. После переноса белков мембрану инкубируют последовательно с антителами, специфичными к исследуемому белку, а затем со вторичными антителами, специфичными к Fc-фрагментам первичных антител, конъюгированными с ферментной (или какой-либо другой) меткой (рис. 1 А). В случае, когда с меткой конъюгированы непосредственно первичные, специфичные к исследуемому антигену антитела, вторичные антитела не требуются (рис. 1 Б). Образовавшиеся в месте локализации исследуемого белка иммунные комплексы «проявляются» с помощью хромогенного субстрата (зависит от типа метки). Чувствительность и специфичность метода сильно зависят от того, какие антитела используются при проведении исследований. Используемые антитела должны быть специфичны к уникальной, характерной только для исследуемого белка последовательности аминокислот. В противном случае возможно взаимодействие (особенно в случае грубых белковых экстрактов) антител с несколькими белковыми молекулами, что в свою очередь приведет к появлению на мембране нескольких окрашенных полос. Идентификация исследуемого белка в таком случае часто бывает затруднительной либо вообще невозможной. Вторым важным фактором, о котором следует помнить при выборе антител, является аффинитет. Чем выше аффинитет используемых антител, тем ярче и четче прокрашиваются белковые полосы, тем выше чувствительность метода. При использовании высокоаффинных антител можно достичь чувствительности 1 нг и даже выше. Для визуализации результата взаимодействия связанного с мембраной антигена и антител используют вторичные антитела, конъюгированные с агентами, способными в определенных условиях давать определенный сигнал. Обычно в качестве такого агента используют фермент (пероксидазу или фосфатазу), продукт реакции которого имеет окраску и выпадает на мембране в виде нерастворимого осадка. Также в данном методе возможно использование флуоресцентных меток.
Электрофорез белков Метод основан на том, что при определённом значении рН и ионной силы раствора белки двигаются в электрическом поле со скоростью, пропорциональной их суммарному заряду. Белки, имеющие суммарный отрицательный заряд, двигаются к аноду (+), а положительно заряженные белки - к катоду (-). Электрофорез проводят на различных носителях: бумаге, крахмальном геле, полиакриламидном геле и др. В отличие от электрофореза на бумаге, где скорость движения белков пропорциональна только их суммарному заряду, в полиакриламидном геле скорость движения белков пропорциональна их молекулярным массам. Разрешающая способность электрофореза в полиакриламидном геле выше, чем на бумаге. Так, при электрофорезе белков сыворотки крови человека на бумаге обнаруживают только 5 главных фракций: альбумины, α1 глобулины, α2 -глобулины, β-глобулины и γ-глобулины (рис. 1-57). Электрофорез тех же белков в полиакриламидном геле позволяет получить до 18 различных фракций. Для обнаружения белковых фракций полоски бумаги или столбики геля обрабатывают красителем (чаще всего бромфеноловым синим или амидовым чёрным). Окрашенный комплекс белков с красителем выявляет расположение различных фракций на носителе. Ионообменная хроматография Так же как и электрофорез, метод основан на разделении белков, различающихся суммарным зарядом при определённых значениях рН и ионной силы раствора. При пропускании раствора белков через хроматографическую колонку, заполненную твёрдым пористым заряженным материалом, часть белков задерживается на нём в результате электростатических взаимодействий. В качестве неподвижной фазы используют ионообменники - полимерные органические вещества, содержащие заряженные функциональные группы. Различают положительно заряженные анионообменники, среди которых наиболее часто используют диэтиламиноэтилцеллюлозу (ДЭАЭ-целлюлозу), содержащую катионные группы, и отрицательно заряженные катионообменники, например карбоксиметилцеллюлозу (КМ-цел-люлозу), содержащую анионные группы.
Выбор ионообменника определяется зарядом выделяемого белка. Так, для выделения отрицательно
Аффинная хроматография. Ядро. Строение и свойства. Ядро клетки — главный центр с генетической информацией, так как в нем находятся хромосомы, содержащие наследственные признаки, закодированные в форме ДНК. Другие носители информации имеют меньшее значение. Положение, форма и размеры ядра могут изменяться, часто параллельно с изменениями интенсивности метаболизма. Ядро чаще всего расположено в центре клетки, и только у растительных клеток с центральной вакуолью — в пристеночной протоплазме. Оно может быть различной формы: · сферическим; · яйцевидным; · чечевицеобразным; · сегментированным (редко); · вытянутым в длину; · веретеновидным, а также иной формы. Диаметр ядра варьирует в пределах от 0,5 мкм (у грибов) до 500 мкм (в некоторых яйцеклетках), в большинстве случаев он меньше 5 мкм. Ядро состоит из: · нуклеоплазмы; · хромосом (хроматина); · ядрышек; · ядерной оболочки, представляющей собой часть эндоплазматического ретикулума. Клеточные ядра образуются только из ядер. Репликация ДНК, т. е. удвоение генетической информации, гарантирует идентичность ядер, несмотря на всю сложность их деления. Главные функции клеточного ядра следующие: · хранение информации; · передача информации в цитоплазму с помощью транскрипции, т. е. синтеза переносящей информацию и-РНК; · передача информации дочерним клеткам при репликации — делении клеток и ядер.
Биосинтез антибиотиков Антибиотики относятся к вторичным метаболитам — низкомолекулярным веществам, функции которых несущественны для роста чистых культур. Вторичные метаболиты (антибиотики, алкалоиды, гормоны роста растений, токсины) производятся представителями ограниченного числа таксономических групп. В отличие от первичных метаболитов, вторичные имеют более сложное химическое строение и образуются, как правило, в ходе длинных метаболических путей. Известно, что образование антибиотиков делает их продуцентов более конкурентоспособными и является мощным фактором в борьбе организмов за существование. Однако это не единственная роль антибиотиков: недавно обнаружено, что данные вещества осуществляют функцию контроля и регуляции процессов метаболизма, выполняя роль специфических эффекторов, контролирующих активность ряда ферментных систем. Кроме этого, существует точка зрения, согласно которой антибиотики могут служить клеточными компонентами, в виде которых клетка избавляется от накопившихся в ней по тем или иным причинам в избытке метаболитов и восстановительных эквивалентов. Биоосинтез большинства вторичных метаболитов, в том числе антибиотиков, обычно осуществляется на определенной стадии развития микроорганизма, в условиях торможения основных процессов анаболизма—биосинтеза нуклеиновых кислот, белка, липидов, полисахаридов. Различают две принципиально различные стадии развития популяции продуцентов антибиотиков. Первая, носящая название трофофаза или фаза сбалансированного роста, характеризуется интенсивным синтезом соединений, требующихся для роста клеток, что сопровождается быстрым потреблением основных питательных компонентов среды. Эта фаза соответствует фазе логарифмического роста периодической культуры микроорганизмов. После исчерпания некоторых ингредиентов среды (в первую очередь, углерода, азота, молекулярного кислорода) и накопления продуктов метаболизма наступает вторая фаза (идиофаза, или фаза несбалансированного роста). Идиофаза отличается заметным торможением или прекращением роста микробной популяции, что соответствует переходу в стационарную фазу роста периодической культуры. Здесь начинают преобладать протеолитические процессы, происходит автолитический распад клеток. Именно в идиофазе многие микроорганизмы начинают синтезировать вторичные метаболиты, к числу которых относятся и антибиотики. Описанные закономерности можно отобразить графически (рис. 18.3).
В большинстве случаев максимум накопления антибиотика в культуральной жидкости наступает после максимума накопления биомассы. Это объясняется тем, что синтез ферментов, ответственных за образование антибиотиков, подавлен (репрессирован) во время трофофазы. Биосинтез антибиотиков — наследственно закрепленное свойство организмов. Гены, ответственные за эти процессы, могут располагаться на хромосомах (нуклеоидах у прокариот), а также на плазмидах. Регуляция процесса синтеза антибиотика осуществляется разными способами: чаще в ходе катаболитной репрессии, с помощью репрессии синтеза ферментов конечным продуктом, а также при аллостерическом ингибировании антибиотиком активности ключевых ферментов своего биосинтеза. При этом важными компонентами среды, оказывающими влияние на биосинтез антибиотиков, являются глюкоза, а также фосфат. Как правило, избыток этих веществ отрицательно сказывается на образовании антибиотиков. Антибиотическая продуктивность организма отражает количество антибиотика (в мкг или в единицах), образованное единицей массы (обычно 1 мг) сухих клеток микроорганизма-продуцента в единицу времени (обычно за 1 ч). Этот параметр зависит, в первую очередь, от вида микроорганизма, фазы роста клеток, состава среды, условий культивирования. Препараты антибиотиков характеризуются разной степенью чистоты и, соответственно, активности, которая выражается в количестве единиц антибиотической активности (ЕД), содержащихся в 1 мг или 1 мл препарата. За единицу антибиотической активности принимают минимальное количество чистого вещества антибиотика, способное подавить развитие определенного числа клеток стандартного штамма микроорганизма (тест-микроба) в единице объема культуральной жидкости. Например, 1 ЕД стрептомицина эквивалентна 1 мкг чистого основания этого вещества, 1 ЕД пенициллина соответствует 0,6 мкг натриевой соли бензилпенициллина. При этом в качестве тест-микроба для стрептомицина используется E.coli, а для пенициллина — Staphylococcus aureus. Синтез биополимеров Схематически все процессы образования полипептидов и полинуклеотидов укладываются в следующую схему:
- где над стрелками обозначены названия соответствующих стадий. Общие принципы реакций Все биополимеры (полипептиды, гетерополисахариды, полинуклеотиды) имеют организацию, - специфическую упаковку в пространстве, поэтому их синтез включает дополнительно и стадию структурирования. А если их звенья еще и отличаются друг от друга, отсюда, чтобы вновь образующийся полимер включал их в строго определенном порядке, необходима матрица, что и отличает генез белков и нуклеиновых кислот от других подобных процессов. Кроме того, любой синтез требует затрат энергии, причем часто в этих случаях используются не обычные макроэрги, а происходит с их помощью предварительная активация субстратов. Исходя из этих предпосылок, можно выделить следующие общие принципы, характерные для этих процессов: 1. Наличие матрицы: в синтезе дочерней ДНК ею служит вся материнская ДНК; для образования РНК используется фрагмент ДНК (ген, или транскриптон); аминокислотная последовательность определяется набором триплетов иРНК. 2. Субстраты предварительно активируются: на первой стадии используются дезоксирибонуклеотидтрифосфаты (дАТФ, дГТФ, дЦТФ, ТТФ), генез РНК требует наличия соответственно АТФ, ГТФ, ЦТФ, УТФ, полипептиды получаются из аминоацил-тРНК. 3. Основные ферменты, обеспечивающие собственно полимеризацию, относятся к классу трансфераз и называются ДНК-, РНК-полимеразы и пептидилтрансфераза. 4. Сложность образования биополимера выделять в этом процессе две фазы: собственно синтез (как бы начерно) и созревание – укладку в пространстве, химическую модификацию. 5. В собственно синтезе выделяют следующие стадии: а) инициацию – начало; б) элонгацию (longus- длинный) – наращивание цепи; в) терминацию – остановку реакций. Репликация ДНК Репликация – синтез дочерней ДНК с использованием в качестве матрицы всей молекулы материнской ДНК. С этого процесса и начинается деление клетки в S-фазу (синтетическую). В основе репликации лежат три принципа: а)комплементарности– азотистые основания образующейся дочерней цепи должны обладать химическим и геометрическим соответствием с подобными веществами материнской нити;полуконсервативности– каждая из синтезированных молекул ДНК состоит из одной материнской и одной дочерней; в)однонаправленности – считывание матрицы идет от 5' к 3' концу. Чтобы порядок азотистых оснований, находящихся внутри нитей ДНК, можно было считывать, необходимо разорвать водородные связи, соединяющие между собой цепи и лишить их спирализации. Для этих целей используются следующие ферменты: топоизомеразаихеликаза. Первая гидролизует на одном из участков полинуклеотида фосфодиэфирная связь, что позволяет этому фрагменту раскрутиться относительно параллельно л а затем восстановить связь. Хеликазаразрушает водородные взаимодействия между противолежащими основаниями участков двух цепей:
Под ее действием фрагменты расходятся, образуя репликативную вилку. Затем с помощьюпраймазы(primer- затравка) синтезируется олигорибонуклеотид – небольшое соединение, состоящее (обратите внимание) из 8-10 монорибонуклеотидов. Оно служит местом, куда крепится основной фермент –ДНК-полимераза. Из-за антипараллельности цепей данные структуры будут локализоваться в разных местах: одна у конца, другая несколько в глубине противоположной нити:
Все дело в том, что ДНК-полимеразыне могут начинать синтез, а способны только добавлять дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся цепи, чем и являетсяпраймер. Описано 3 класса данных энзимов: в ядрах присутствуют полимеразы альфа (Роlα), ответственные за хромосомную репликацию, полимеразы бета (Роlβ), которые используются при необходимости при репарации. Третий их представитель – ДНК-полимераза гамма (Роlγ) осуществляет синтез кольцевого генома митохондрий. Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом макроэргической связи в дНТФ и отщеплением пирофосфата. Важная деталь – генетический материал живых организмов имеет огромные размеры, но реплицируется с высокой точностью. В среднем при воспроизведении генома (ДНК длиной более 3 млрд пар нуклеотидов) возникает не более 3-х ошибок благодаря наличию специальных механизмов, осуществляющих необходимую коррекцию. Суть последней в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй – перед присоединением нового нуклеотида. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний мономер удлиняющейся дочерней нити ДНК образовал правильную (комплементарную) пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии произошло ошибочное спаривание оснований, то фермент возвращает последнее добавленное звено, после чего освободившееся место занимает правильный нуклеотид. Параллельно с ростом цепи продолжается деспирализация и разделение цепей с помощью топоизомеразы и хеликазы, удлинение репликативной вилки. А так как процесс осуществляется лишь в одном (5' – 3') направлении, что позволяет происходить этому явлению непрерывно только на одной из матричных нитей. На антипараллельной синтез ограничивается сравнительно короткими фрагментами (100-1000 мононуклеотидов), названными по имени обнаружившего их ученого фрагментами Оказаки(с постоянным предварительным синтезом праймеров):
Так продолжается до тех пор, пока вся матрица не выполнит свою функцию, причем во время репликации специальные нуклеазы вычленяют праймеры. Если позволяет направление, ДНК-полимеразы застраивают возникшие бреши, а сшивание сближенных фрагментов обеспечивается ДНК-лигазами– так завершается собственно синтез (стадия терминации). В тех концах дочерних цепей, где находились праймеры, которые позднее были удалены, не происходит достраивания дезоксирибонуклеотидами (невозможно считывание в обратном направлении), отсюда при каждом делении клетки молекулы новых цепей укорачиваются на 10-20 нуклеотидов, но объем информации при этом не уменьшается, так как потерянные участки ее не несли. Дело в том, что когда в эмбриональных тканях шли усиленные митозы параллельно с этим осуществлялись обратные транскрипции. С помощьютеломеразы концы ДНК наращивались теломерными повторами ТТАGGG, роль матрицы при этом выполняла РНК, включающая 450 мононуклеотидов. Поскольку многократно повторенные теломерные последовательности не являются кодирующими, их утрата в процессе деления не приводит к потере информативных участков, тем самым обеспечивается передача генетического материала в поколениях клеток. Однако в родившемся организме активность теломераз регистрируется лишь в половых, раковых, стволовых клетках, в соматических же эти ферменты не работают. Теломеры постепенно укорачиваются, что в конце концов запускает процессы остановки клеточного цикла и провоцирует апоптоз. Интересный факт: ученые, исследующие рак, до сих пор используют культуру клеток Неlа из опухоли женщины по имени Генриетта Лакс. Эта больная умерла в 1951 году, но клетки ее новообразования продолжают делиться и собираются жить вечно. Происходящее в момент деления клетки удлинение синтезирующихся нитей ДНК сопровождается их созреванием, что включает спирализацию, суперспирализацию (закручивание цепей вокруг гистоновых нуклеосом), химическую модификацию (гидроксилирование, метилирование, гидрирование азотистых оснований) с образованием минорных оснований, соединение с помощью ионных связей с белками, катионами металлов (К+, Са++, Мg++, Мn++, Fе++, Сu++и т.д.), которые стабилизируют или, наоборот, дестабилизируют вновь синтезированную молекулу. Синтез и процессинг РНК В отличии от ДНК, синтез которой происходит в момент деления клетки и служит источником генетической информации для последующих поколений, различные РНК необходимы для генерирования молекул белков. Процесс синтеза рибонуклеиновых кислот с использованием в качестве матрицы фрагментов ДНК называют транскрипцией. Все гены в хромосоме (ДНК с белками) в отсутствии факторов транскрипции находится в выключенном (репрессированном) состоянии, потому что нуклеосомы блокируют области инициирования каждого промотора. Ген будет транскрибироваться после вытеснения нуклеосомы и связывания. Та из двух цепей ДНК, на которой пойдет этот процесс, называется кодирующей. Инициация включает раскручивание участка, разрыв водородных связей, что раскрывает азотистые основания транскриптона, в котором выделяют промотор, собственно ген (единицу транскрипции), терминатор. Первая функциональная единица (промотор) служит для с ним РНК-полимеразы, которая объединяет рибонуклеотиды в последовательность, комплементарную кодирующей цепи гена. Момент терминации распознается терминирующим белком – р-фактором. Интересно, что в районе промотора расположены сигнальные последовательности двух типов. Одна из них указывает, где должна начаться транскрипция, а другая определяет, как часто должно происходить это событие. Первый локус называют ТАТА-бокс, второй – СААТ-бокс. Кроме них регуляторная зона включает энхансеры или сайленсеры, изменяющие скорость транскрипции, а также гормончувствительные элементы (ГЧЭ), после взаимодействия с которыми кортикостероиды, андрогены, эстрогены, Т3, сАМФ регулируют экспрессию генов. Практически все первичные транскрипты РНК у эукариот подвергаются сложному процесссингу, т.е. созреванию, начинающемуся в ядре. Будущая иРНК получается после копирования, реакций расщепления, вычленения, легирования, включения дополнительных адениловых нуклеотидов. Ведь свежеполученная РНК содержит неинформативные вставки, считанные с интронов генов, т.е. представляют чередование траслируемых и нетранслируемых участков (гетерогенная ядерная РНК). Процесс удаления интронов и последующего сшивания экзонов получил названиесплайсинг (от англ.tosplice– соединять концы, сшивать). Он требует наличия специальных ферментов, поэтому осуществляется в крупном рибонуклеопротеидном комплексе – тельце, содержащем РНК и белки, необходимые для эффективного сплайсинга. Эта структура называетсясплайсосома. Кроме того к 5'-концу г/яРНК присоединяется 7-метилгуанозинфосфат (кэпирование), который позднее служит стартовой точкой синтеза белка, к 3'-концу надстраивается полиаденилатный хвост (около 200 мононуклеотидов), защищающий иРНК от действия нуклеаз. Параллельно иРНК уже вышедшая из ядра, плотно упаковывается в пространстве. Первичный транскрипт рРНК не содержит интронов, интенсивно метилируется и расщепляется в ядрышке специфическими РНК-азами на 5S, 18S, 28-рРНК, затем они связываются с белками, образуя рибосомы. Подобным образом (путем частичного гидролиза) осуществляется из первичных транскриптов образование тРНК, формирование вторичной, третичной структуры, химическая модификация. Критическое значение имеет этап сплайсинга в области антикодона, т.к. от него зависит точность выполнения адапторной функции при синтезе белка. Генерирование полипептидов Образно говоря, для осуществления двух предыдущих стадий использовался один язык – нуклеотидный, третья же стадия – синтез полипептидов – говорит на другом языке – языке аминокислот. Следовательно, требуется перевод. Поэтому данный процесс и называется трансляцией. Это строительство аминокислотной последовательности на иРНК в качестве матрицы. Для успешного перевода сведений, записанных в генах ДНК, в строго специфический порядок аминокислот в белке используется биологический (генетический) код. Биологические наноструктуры Saccharomycescerevisiae Дрожжи Saccharomyces cerevisiae — это непатогенные одноклеточные микроорганизмы с диаметром клетки примерно 5 мкм, которые во многих отношениях представляют собой эукариотический аналогE. coli. Их генетика, молекулярная биология и метаболизм детально изучены.S. cerevisiae размножаются почкованием и хорошо растут на такой же простой среде, как иЕ. coli. Их способность к превращению сахара в этанол и углекислый газ издавна использовалась для изготовления алкогольных напитков и хлеба. В настоящее время ежегодно во всем мире расходуется более 1 млн. тонн S. cerevisiae. ДрожжиS. cerevisiae представляют также большой научный интерес. В частности, они являются наиболее удобной моделью для исследования других эукариот, в том числе человека, поскольку многие гены, ответственные за регуляцию клеточного деленияS. cerevisiae, сходны с таковыми у человека. Это открытие способствовало идентификации и характеристике генов человека, отвечающих за развитие новообразований. Широко используемая генетическая система дрожжей (искусственная хромосома) является непременным участником всех исследований по изучению ДНК человека. В 19% г. была определена полная нуклеотидная последовательность всего набора хромосомS. cerevisiae, что еще более повысило ценность этого микроорганизма для научных исследований. Такая работа на эукариотах была выполнена впервые. Синтезированный бактериальной клеткой эукариотический белок часто приходится подвергать ферментативной модификации, присоединяя к белковой молекуле низкомолекулярные соединения — во многих случаях это необходимо для правильного функционирования белка. К сожалению, E. coli и другие прокариоты не способны осуществлять эти модификации, поэтому для получения полноценных эукариотических белков используютS. cerevisiae, а также другие виды дрожжей:Kluyveromyces lactis, Saccharomyces diastaticus, Schizisaccharomyces pombe,Yarrowia lipolytica, Pichia postons, Hansenula pofymorpha. Наиболее эффективными продуцентами полноценных рекомбинантных белков являютсяP. pastoris и H. polymorpha. Escherichia coli – один из наиболее изученных организмов. За последние пятьдесят лет удалось получить исчерпывающую информацию о генетике, молекулярной биологии, биохимии, физиологии и общей биологии Escherichia coli. Это грамотрицательная, подвижная полочка длиной менее 10 мкм. Средой ее обитания является кишечник человека и животных, но она также может обитать в почве и в воде. Обычно, кишечная палочка не патогенна, но при определенных условиях может вызывать заболевание человека и животных. Благодаря способности размножаться простым делением на средах, содержащих только ионы Na, K+, Mg2+, Ca2+,NH4+, Cl-, HPO42- и SO42-, микроэлементы и источник углерода (например, глюкозу), E.coli стала излюбленным объектом научных исследований. Эшерихий - факультативные анаэробы, хорошо растут на обычных простых и синтетических питательных средах при температуре от 15 до 46°С. Оптимальная температура для роста - 37-38°С. Хорошо растут при рН среды, близком к нейтральной реакции (7,2-7,4). На плотных питательных средахЕ. coliобразуют круглые выпуклые колонии средней величины, влажные, с гладкой блестящей поверхностью с ровным краем (S-форма) или плоские, сухие со слегка волнистым краем и шероховатой поверхностью (R-форма). В жидких средах эшерихий растут в виде интенсивного равномерного помутнения среды, образуя осадок, иногда пленку на поверхности или кольца на стенке пробирки. Осадок разбивается при встряхивании, образуя гомогенную взвесь. На селективно-дифференциальной среде Эндо лактозоположительные штаммы образуют колонии малиново-красного цвета с металлическим блеском или без него. На агаре Левина (среда с эозином и метиленовым синим) - колонии темно-фиолетового цвета. Бактерии, не ферментирующие или замедленно ферментирующие лактозу, на среде Эндо формируют сероватые колонии и светло-розовые - на агаре Левина. На Сорбитол агаре абсолютное большинство эшерихий образуют колонии красного цвета как на среде Эндо. Исключением являются бактерии серовара О157:Н7, не изменяющие цвет среды и образующие сероватые колонии. Модификация хроматина Комплекс белков с ядерной ДНК клеток называют хроматином.
Гистоны - белки с молекулярной массой 11-21 кД, содержащие много остатков аргинина и лизина. Благодаря положительному заряду гистоны образуют ионные связи с отрицательно заряженными фосфатными группами, расположенными на внешней стороне двойной спирали ДНК. Существует 5 типов гистонов. Четыре гистона Н2А, Н2В, НЗ и Н4 образуют октамерный белковый комплекс (Н2А, Н2В, НЗ, Н4)2, который называют "нуклеосомный кор" (от англ. nucleosome core). Молекула ДНК "накручивается" на поверхность гистонового октамера, совершая 1,75 оборота (около 146 пар нуклеотидов). Такой комплекс гистоновых белков с ДНК служит основной структурной единицей хроматина, её называют "нуклеосома". ДНК, связывающую нуклеосомные частицы, называют линкерной ДНК. В среднем линкерная ДНК составляет 60 пар нуклеотидных остатков. Молекулы гистона H1 связываются с ДНК в межнуклеосомных участках (линкерных последовательностях) и защищают эти участки от действия нуклеаз (рис. 4-8). Рис. 4-8. Структура нуклеосом. Восемь молекул гистонов (Н2А, Н2В, НЗ, Н4)2 составляют ядро нуклеосомы, вокруг которого ДНК образует примерно 1,75 витка.
Строение нуклеиновых кислот. Подобно белкам, нуклеиновые кислоты — биополимеры, а их функция заключается в хранении, реализации и передаче генетической (наследственной) информации в живых организмах. Существует два типа нуклеиновых кислот — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Мономерами в нуклеиновых кислотах служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза — в ДНК, рибоза — в РНК) и остаток фосфорной кислоты. В ДНК входят четыре вида нуклеотидов, отличающихся по азотистому основанию в их составе, — аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т). В молекуле РНК также имеется 4 вида нуклеотидов с одним из азотистых оснований — аденином, гуанином, цитозином и урацилом (У). Таким образом, ДНК и РНК различаются как по содержанию сахара в нуклеотидах, так и по одному из азотистых оснований (табл. 1).
Состав нуклеиновых кислот Нуклеиновые кислоты представляют собой полимеры, построенные из нуклеотидов, соединенных между собой фосфодиэфирными связями. Каждый нуклеотид состоит из остатков азотистого основания, пентозы и фосфорной кислоты. Различают пиримидиновые и пуриновые основания, называемые также соответственно пиримидины и пурины. Пиримидиновые основания являются производными пиримидина:
пуриновые основания – производными пурина: К пиримидинам относятся урацил, тимин и цитозин, к пуринам – аденин и гуанин: В состав ДНК входят тимин, цитозин, аденин и гуанин, в состав РНК – те же основания, только вместо тимина входит урацил. Кроме азотистых оснований, нуклеиновые кислоты содержат пентозы: ДНК – D-дезоксирибозу, а РНК – D-рибозу. Углеводы находятся в виде b-аномера фуранозной формы:
Азотистое основание связывается с углеводом за счет гликозидного гидроксила. Образуется нуклеозид. Схематически образование нуклеозида можно изобразить так:
В состав нуклеиновых кислот входят 8 нуклеозидов, 4 – в состав РНК и 4 – в состав ДНК (рис. 39). Нуклеозиды, входящие в состав РНК:
Нуклеозиды, входящие в состав ДНК:
Рис. 39. Нуклеозиды
Нуклеозид, связанный с остатком фосфорной кислоты, называется нуклеотидом:
При этом остаток фосфорной кислоты может быть связан с 3’- или 5’- атомом углерода:
Сокращенно аденозин-5’-монофосфат обозначается как АМФ. Если нуклеотид образован дезоксорибозой, аденином и одним остатком фосфорной кислоты, то он будет носить название дезоксиаденозинмонофосфат, или сокращенно дАМФ. В таблице 5 представлена номенклатура нуклеотидов.
2. Назовите функции ДНК
Выделяют три основные функции ДНК. Хранение наследственной информации. Порядок нуклеотидов определяет первичную структуру белков. Первичная структура, в свою очередь, обуславливает свойства белков, а следовательно, особенности строения и функционирования клеток. Таким образом, в ДНК закодирована информация обо всех свойствах клеток, тканей и органов. Участок молекулы ДНК, кодирующий первичную структуру одной белковой цепи, называют геном. Передача наследственной информации следующему поколению клеток. Эта функция осуществляется благодаря способности ДНК к удвоению (редупликации). После деления в каждую дочернюю клетку попадает одна из двух идентичных молекул ДНК, являющихся точной копией материнской ДНК. Передача наследственной информации из ядра в цитоплазму. Почти вся ДНК находится в ядре; синтез же белка происходит в цитоплазме клетки. Соответственно, необходим посредник, передающий описание первичной структуры белка от ДНК к рибосоме. В роли такого посредника выступает информационная РНК, которая синтезируется на одной из цепей ДНК, копируя по принципу комплементарности последовательность нуклеотидов определенного гена.
3. В чём различаются строение ДНК и РНК.
|
Последнее изменение этой страницы: 2019-04-19; Просмотров: 483; Нарушение авторского права страницы

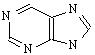
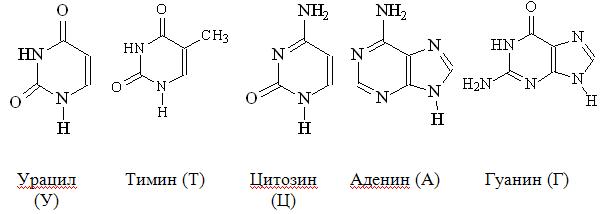
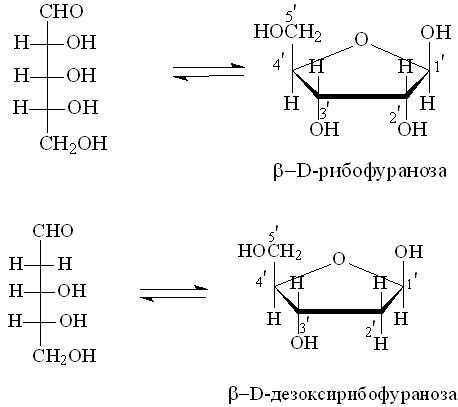
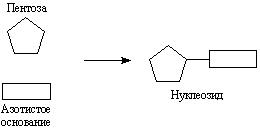
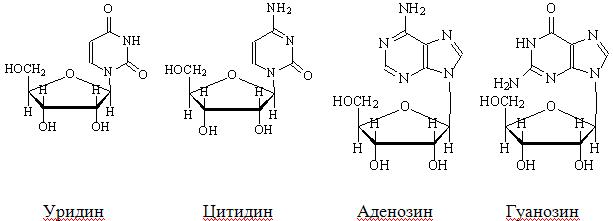
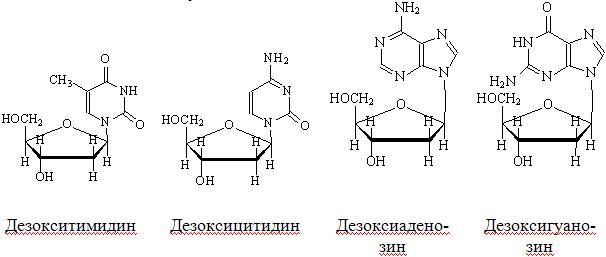





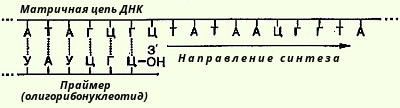
 фибрин.
фибрин.
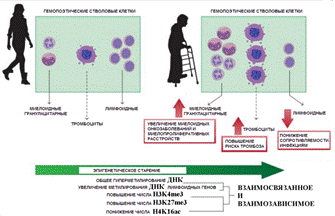




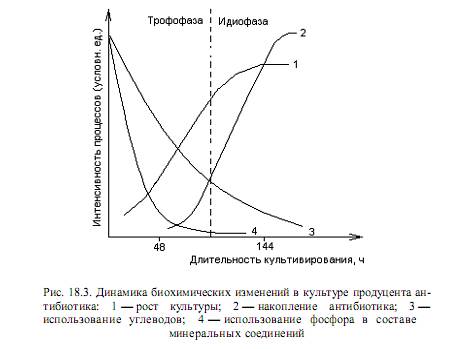

 ежащего:
ежащего: