|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
С труктура и функции белков
Белки (протеины) составляют 50% от сухой массы живых организмов.
Белки состоят из аминокислот. У каждой аминокислоты есть аминогруппа и кислотная (карбоксильная) группа, при взаимодействии которых получается пептидная связь, поэтому белки еще называют полипептидами. Структуры белка Первичная – цепочка из аминокислот, связанных пептидной связью (сильной, ковалентной). Чередуя 20 аминокислот в разном порядке, можно получать миллионы разных белков. Если поменять в цепочке хотя бы одну аминокислоту, строение и функции белка изменятся, поэтому первичная структура считается самой главной в белке.
Вторичная – спираль. Удерживается водородными связями (слабыми).
Третичная – глобула (шарик). Четыре типа связей: дисульфидная (серный мостик) сильная, остальные три (ионные, гидрофобные, водородные) – слабые. Форма глобулы у каждого белка своя, от нее зависят функции. При денатурации форма глобулы меняется, и это сказывается на работе белка.
Четвертичная – имеется не у всех белков. Состоит из нескольких глобул, соединенных между собой теми же связями, что и в третичной структуре. (Например, гемоглобин.) Функции белков: 1) защитная (интерферон усиленно синтезируется в организме при вирусной инфекции); 2) структурная (коллаген входит в состав тканей, участвует в образовании рубца); 3) двигательная (миозин участвует в сокращении мышц); 4) запасная (альбумины яйца); 5) транспортная (гемоглобин эритроцитов переносит питательные вещества и продукты обмена); 6) рецепторная (белки-рецепторы обеспечивают узнавание клеткой веществ и других клеток); 7) регуляторная (регуляторные белки определяют активность генов); 8) белки-гормоны участвуют в гуморальной регуляции (инсулин регулирует уровень сахара в крови); 9) белки-ферменты катализируют все химические реакции в организме; 10) энергетическая (при распаде 1 г белка выделяется 17 кдж энергии). Биосинтез белка Основываясь на данных биспиральной антипараллельной структуры и химического состава ДНК (см. Химия нуклеиновых кислот), а также на данных о значении "активированной" формы энергии для биосинтеза полимерных молекул, А. Корнберг еще в 1955 г. впервые указал на возможность синтеза энзиматическим путем ДНК в бесклеточной системе в присутствии изолированной из Е. coli ДНК-полимеразы и предшественников дезоксирибонуклеозидтрифосфатов; реакция сводится к синтезу новой молекулы ДНК:
Сущность реакции сводится к присоединению мононуклеотидных единиц к свободному 3'-гидроксильному концу ДНК и образованию новой полинуклеотидной цепи в направлении 5'-> 3'; схематически она может быть представлена в виде:
Реакция требует присутствия одноцепочечной ДНК или в крайнем случае небольших фрагментов полинуклеотидов. Пока еще нет полной ясности о значении предобразованной ДНК в действии ДНК-полимераз. Более вероятно мнение, что ДНК служит матрицей, на которой фермент комплементарно и антипараллельно синтезирует новую цепь ДНК. Это можно представить в виде схемы:
Как видно из схемы, последовательность вновь синтезируемого полинуклеотида отражает первичную структуру, и обе цепи по отношению друг к другу являются антипараллельными. Следует указать, что ДНК-полимераза I оказалась наиболее активной в тех случаях, когда добавленная в качестве матрицы молекула ДНК была предварительно денатурирована. В этих случаях соблюдалась не только точная последовательность оснований, но и ряд физических свойств исходной ДНК. Напротив, если в опытах in vitro была добавлена нативная двухцепочечная ДНК, то вновь синтезированный полинуклеотид значительно отличался от ДНК и, в частности, не обладал свойственной ДНК биологической активностью.
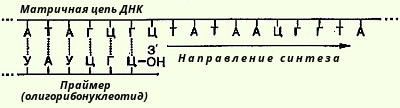
Были предприняты другие подходы к выяснению механизма полимеразной реакции. В лаборатории Корнберга был открыт фаг (ФХ174), содержащий одноцепочечную кольцевую ДНК; когда эта молекула была использована в качестве матрицы в ДНК-полимеразной реакции, то в этих условиях была получена биологически активная ДНК фага. В этой работе важную роль сыграл другой фермент, названный ДНК-лигазой, обладающий способностью катализировать репарацию ДНК, соединение концов разрывов в молекуле ДНК. Было показано, что в процессе репликации одноцепочечная ДНК фага ФХ174 проходит стадию образования двухцепочечной кольцевой ДНК. Применив ряд остроумных подходов, Корнберг и сотр. в опытах in vitro создали искусственную молекулу фага ФХ174, обладающую способностью поражать (инфицировать) Е. coli, вызывая лизис бактерии. Последовательность событий в этой работе может быть представлена в следующей схеме, где исходную молекулу кольцевой ДНК фага ФХ174 пометим плюсом (+), а вновь синтезируемую молекулу-минусом (-) (рис. 106). В исследованиях Меселсона и Сталя получила подтверждение гипотеза полуконсервативного механизма синтеза ДНК в клетках, означающего наличие в каждой из двух дочерних молекул одной из родительских цепей ДНК (рис. 107). Репликация ДНК, как правило, начинается с какой-нибудь одной точки на обеих цепях, и направление роста каждой цепи также идет в одном направлении. Кроме того, как было указано выше, ДНК-полимераза I не реплицирует нативную двухцепочечную ДНК и осуществляет синтез только в одном направлении, а именно в направлении 5'->3'. Была предложена интересная гипотеза, объясняющая репликацию обеих цепей нативной молекулы ДНК. Этот механизм требует наличия трех ферментов: ДНК-полимеразы, ДНК-лигазы и эндонуклеазы. Репликация начинается с разрыва в одной из двух цепей под действием эндонуклеазы. Затем к этому месту присоединяется ДНК-полимераза и начинается непрерывный синтез нового олигонуклеотида на одной из двух родительских цепей в направлении 5'->3', обратный направлению исходной цепи ДНК. Из второй цепи родительской ДНК идет синтез прерывистый, сопровождающийся образованием фрагментов ДНК, также в направлении 5'->3' с последующим объединением фрагментов ДНК при участии ДНК-лигаз в единую полинуклеотидную молекулу. Подобный механизм челночного синтеза ДНК легко объясняет фактические данные накопления коротких фрагментов ДНК у Е. coli во время репликации ДНК. Совсем недавно было показано, что инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на материнской ДНК необычного затравочного олигорибонуклеотида, названного праймером, со свободной гидроксильной группой, у 3'-углеродного атома рибозы. Этот олигорибонуклеотид, содержащий около 50 нуклеотидных остатков, синтезируется комплементарно на матрице ДНК при участии особого фермента - РНК-полимеразы (см. ниже). Предполагается, что именно с этой точки концевого 3'-гидроксила рибозы праймера начинается истинный синтез дочерней цепи ДНК, комплементарной родительской. В дальнейшем этот фрагмент РНК, ковалентно присоединенный к новообразованной цепи ДНК, разрушается под действием нуклеаз и возникшая брешь застраивается олигодезоксирибонуклеотидом при помощи той же ДНК-полимеразы. Вполне допустимо предположение, что синтез праймера изолигорибонуклеотида имеет биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
Классификация белков |
Последнее изменение этой страницы: 2019-04-19; Просмотров: 214; Нарушение авторского права страницы