|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ДИФФУЗИОННАЯ СПОСОБНОСТЬ ЛЕГКИХ. ЛЕГОЧНАЯ ВЕНТИЛЯЦИЯСтр 1 из 7Следующая ⇒
ЛЕГОЧНАЯ ВЕНТИЛЯЦИЯ Легочная вентиляция, или просто дыхание, представляет собой процесс перемещения воздуха в легкие и из легких. Анатомическая структура респираторной системы показана на рис. 9.1. Воздух, как правило, поступает в легкие через нос; рот используется в том случае, когда потребность в воздухе превышает количество, которое может попасть в легкие через нос. Дыхание через нос предпочтительнее, поскольку воздух согревается и увлажняется, проходя через пространство неправильной формы (носовую раковину). Кроме того, при прохождении через носовую раковину пыль и другие частицы оседают на слизистой оболочке носоглотки. Таким образом осуществляется фильтрация всех частиц, за исключением мельчайших, что снижает раздражение и вероятность респираторных инфекций. Из носа и рта воздух движется по глотке, гортани, трахее, бронхам и бронхиолам, пока не 174
достигнет самых маленьких респираторных единиц — альвеол. Альвеолы являются местом газообмена в легких. Дыхание через нос обеспечивает согревание и увлажнение воздуха, а также фильтрацию инородных частиц Легкие не прикреплены непосредственно к ребрам, а " подвешены" с помощью плевральных полостей, окутывающих легкие. Они содержат тонкий слой плевральной жидкости, которая снижает трение при дыхательных движениях. Кроме
Рис. 9.1. Анатомия дыхательной системы того, эти полости соединены с легкими, а также с внутренней поверхностью грудной клетки, которая придает легким свой размер и форму по мере ее расширения и сокращения. Взаимосвязь между легкими, плевральными полостями и грудной клеткой определяет потоки вдыхаемого и выдыхаемого воздуха. Рассмотрим обе фазы —вдох и выдох. ВДОХ Вдох — активный процесс, в котором участвуют диафрагма и внешние межреберные мышцы. Динамика выполнения вдоха показана на рис. 9.2. Движение ребер и грудины осуществляется внешними межреберными мышцами. Ребра движутся вверх и в стороны подобно ручке ведра. Движения грудины направлены вверх и вперед и напоминают движения ручки насоса. Что касается диафрагмы, то она сокращается, опускаясь вниз к брюшной полости. Эти действия вызывают увеличение объема грудной клетки и легких. При расширении легких, воздух, находящийся в них, заполняет больше пространства и давление в легких снижается (рис. 9.2, в). В результате давление в легких (внутрилегочное давление) становится меньшим, чем давление окружающего воздуха. Поскольку дыхательные пути открыты, воздух устремляется в легкие, чтобы снизить разность давления. Таким образом при вдохе в легкие попадает воздух. В условиях выполнения значительной физической нагрузки осуществлению вдоха способствуют другие мышцы: лестничные (передняя, средняя и задняя) и грудино-ключично-сосцевидная, расположенные в области шеи, а также грудные. С их помощью ребра поднимаются выше, чем при обычном дыхании. Изменение давления, обеспечивающее адекватное вентилирование в состоянии покоя, очень незначительно. Например, при обычном атмосферном давлении (760мм рт.ст.) в результате вдоха давление в легких (внутрилегочное давление) может снизиться всего на 3 мм рт.ст. Однако при максимальном респираторном усилии, например, во время выполнения значительной физической нагрузки, внутрилегочное давление может снизиться на 80— 100 мм рт.ст.! ВЫДОХ В состоянии покоя выдох, как правило, — пассивный процесс, который включает расслабление дыхательных мышц и эластическую тягу легочной ткани (рис. 9.3, а). При расслаблении диафрагмы она принимает свое обычное дугообразное 175 Во время вдоха
Ребра движутся подобно Грудина движется вперед и ручке ведра, расширяя объемы груди вверх подобно ручке насоса, расширяя объемы груди (по бокам (спереди и сзади) (вид сбоку )
)
Мышцы сокращаются, легкие Вдох: воздух устремляется внутрь, уравновешивая давление расширяются: Давление в легких< атмосферного давление
Рис. 9.2. Процесс выполнения вдоха: а — объем грудной клетки в покое; б — увеличение объема при мышечном сокращении; в — увеличение обеспечивает снижение давления в легких и устремление воздуха в них положение. В результате расслабления внешних межреберных мышц ребра и грудина опускаются вниз, занимая обычные для состояния покоя положения. Как только это происходит, легочная ткань также принимает положение, характерное для состояния покоя. Как следует из рис. 9.3, 6, это приводит к повышению давления в грудной клетке, вследствие чего из легких выходит воздух. Выдох завершен. При дыхании с усилием выдох становится более активным процессом. Внутренние межреберные мышцы более активно тянут ребра вниз. Им могут помогать широчайшая мышца спины и поясничная квадратная мышца. Сокращение мышц живота повышает внутрибрюшное давление, вызывая движение внутренних органов вверх к диафрагме и ускоряя ее возврат в исходное дугообразное положение. Эти мышцы, кроме того, тянут грудную клетку вниз и вовнутрь. 176
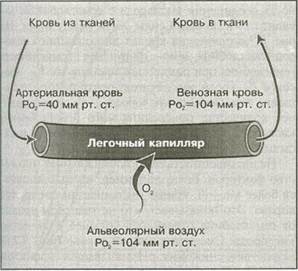
Рис. 9.3. Процесс выполнения выдоха: а — объем грудной клетки возвращается к объему, характерному для состояния покоя; б — воздух устремляется из легких наружу Изменение внутрибрюшного и внутригрудно-го давления не только способствует выполнению дыхания с усилием, но и обеспечивает возврат венозной крови в сердце. С увеличением давления оно передается в большие вены, по которым кровь транспортируется к сердцу через брюшной и грудной участки. При снижении давления размер вен возвращается к исходному и они наполняются кровью. Изменение давления в брюшном и грудном участках вынуждает кровь идти в вены, обеспечивая ее возврат с помощью " выдаивающего" действия. В этом заключается сущность венозного возврата. Сокращения мышц при выполнении физической нагрузки также обеспечивают подобное " выдаивающее " действие, способствуя венозному возврату 2) выводит диоксид углерода из венозной крови. Для осуществления диффузии необходимы воздух, транспортирующий кислород в легкие, и кровь, утилизирующая этот кислород и выделяющая диоксид углерода. Воздух поступает в легкие во время легочной вентиляции, при этом осуществляется газообмен между ним и кровью. В ОБЗОРЕ... 1. Легочная вентиляция (дыхание) представляет собой процесс, обеспечивающий поступление воздуха в легкие и выведение его из них. Она состоит из двух фаз — вдоха и выдоха. 2. Вдох — активный процесс, во время которого увеличиваются размеры диафрагмы и внешних межреберных мышц, а следовательно, и объем грудной клетки. Это приводит к снижению давления в легких и поступлению в них воздуха. 3. Выдох, как правило, — пассивный процесс. Дыхательные мышцы расслабляются, а эластичная ткань легких принимает исходное положение, обеспечивая возвращение объема грудной клетки в обычное исходное положение. Это приводит к увеличению давления в легких и выделению из них воздуха. 4. Вдох и выдох, выполняемые с усилием, — активные процессы, зависящие от мышечных сокращений. Кровь из большинства участков тела возвращается по полой вене в легочную (правую) часть сердца. Из правого желудочка она выкачивается через легочную артерию в легкие, прокладывая 177 себе путь к легочным капиллярам. Эти капилляры образуют плотную сеть вокруг альвеолярных мешочков. Легочные капилляры настолько малы, что эритроциты могут передвигаться по ним лишь в один ряд, подвергаясь воздействию окружающей легочной ткани. Именно здесь происходит процесс диффузии. ЛЕГОЧНАЯ МЕМБРАНА Газообмен между воздухом в альвеолах и кровью в легочных капиллярах осуществляется через легочную мембрану (или как ее еще называют, — альвеолярно-капиллярную мембрану). Эта мембрана (рис. 9.4) состоит из альвеолярной стенки; капиллярной стенки и их базальных мембран. Респираторная мембрана очень тонкая — 0.5 — 4.0 мкм. Вследствие этого приблизительно 300 млн альвеол находятся в непосредственной близости от циркулирующей по капиллярам крови. Тем не менее эта мембрана является значительным препятствием для осуществления газообмена. Рассмотрим, как он происходит. ПАРЦИАЛЬНОЕ ДАВЛЕНИЕ ГАЗОВ Воздух, которым мы дышим, представляет собой смесь газов. Каждый из них оказывает давление, пропорциональное его концентрации в этой смеси. Давление каждого газа в смеси называется парциальным давлением. Согласно закону Дальтона, общее давление смеси газов равно сумме парциальных давлений каждого газа в этой смеси. Рассмотрим состав воздуха, которым мы дышим. Он состоит на 79, 04 % из азота (М^), на Обмен кислорода ^ воздуха при стандартном атмосферном давлении равно 159 мм рт.ст. Однако во время вдоха в альвеолах оно снижается до 100 — 105 мм рт.ст. Поступивший воздух смешивается с тем, который находится в альвеолах; альвеолярный воздух содержит большее количество водяного пара и диоксида углерода, обеспечивающих общее давление. Свежий воздух, вентилирующий легкие, постоянно смешивается с воздухом, остающимся в альвеолах, тогда как некоторые из альвеолярных газов выводятся во внешнюю среду. В результате этого концентрации альвеолярного газа остаются относительно постоянными. Кровь, лишенная большей части кислорода, поступает в легочные капилляры с порядка Р. 40 — 45 мм рт.ст. (рис. 9.5). Это почти на 55 — 65 мм рт.ст. меньше, чем Р<.^ в альвеолах. Иными словами, градиент давления кислорода через легочную мембрану составляет, как правило, 55 — 65 мм рт.ст. Как отмечалось выше, этот градиент давления содействует перемещению кислорода из альвеол в кровь, чтобы уравновесить давление кислорода по обе стороны легочной мембраны. Р»^ в альвеолах относительно постоянно и составляет около 104 мм рт.ст. На артериальном конце капилляра в момент начала газообмена ^о, в крови составляет всего 40 мм рт.ст., однако по мере продвижения крови по капилляру газообмен протекает более интенсивно. К моменту достижения венозного конца капилляра парциаль
Рис. 9.5. Парциальное давление кислорода в крови, проходящей через легочный капилляр ное давление кислорода в крови соответствует давлению в альвеоле. Ро2 довольно быстро уравновешивается и его показатели в альвеолах и в крови достигают 104 мм рт.ст. Следовательно, кровь, покидающая легкие через легочные вены, возвращается в сердце с достаточным количеством кислорода, который может быть использован тканями. Интенсивность диффундирования кислорода из альвеол в кровь называется диффузионной способностью кислорода. В состоянии покоя на каждый 1 мм рт.ст. разности давления каждую минуту в кровь диффундирует около 23 мл кислорода. При максимальном мышечном усилии потребление кислорода у нетренированных людей может повышаться до 45 мл-кг -мин, а у сильнейших спортсменов, занимающихся циклическими видами спорта, — до 80 мл-кг^-мин" '. Повышение диффузионной способности кислорода при переходе от состояния покоя к выполнению физической нагрузки обусловлено относительно неэффективной, " вялой" циркуляцией крови в легких, в основном обусловленной ограниченной перфузи-ей верхних участков легких вследствие действия силы тяжести. В то же время при максимальном усилии кровоток в легких усиливается главным образом вследствие повышения давления крови, обусловливающего более интенсивную перфузию легких. Т Чем выше градиент давления через легочную мембрану, тем быстрее диффузия кислорода через нее У спортсменов с более высокими аэробными возможностями, как правило, выше и диффузионная способность кислорода. По-видимому, это обусловлено увеличенным сердечным выбросом, большей альвеолярной площадью и пониженным сопротивлением диффузии через легочную мембрану. Обмен диоксида углерода Обмен диоксида углерода, как и обмен кислорода, зависит от градиента давления. Как видно из рис. 9.6, парциальное давление углекислого газа (рсо^ ) в крови, проходящей по альвеолам, составляет около 45 мм рт.ст. В альвеолах оно составляет около 40 мм рт.ст. Несмотря на относительно небольшой градиент давления (около 5 мм рт.ст.), он более чем достаточен. Растворимость диоксида углерода в легочной мембране в 20 раз больше, чем растворимость кислорода, поэтому СО^ диффундирует через нее намного быстрее. Показатели парциального давления газов, участвующих в процессе диффузии в легких, приводятся в табл. 9.1. ! 2* 179
Рис. 9.6. Парциальное давление диоксида углерода в крови, проходящей через легочной капилляр В ОБЗОРЕ... 1. Диффузия представляет собой процесс газообмена через легочную мембрану в альвеолах. 2. Величина газообмена через легочную мембрану главным образом зависит от парциального давления каждого из газов. В то же время определенное значение имеют растворимость газов и температура. Газы диффундируют из области с более высоким парциальным давлением данного газа в область с более низким его давлением. Таким образом, кислород поступает в кровь, а диоксид углерода выводится из нее. 3. Диффузионная способность кислорода повышается при переходе из состояния покоя в состояние выполнения физической нагрузки. Когда организму требуется больше кислорода, процесс газообмена интенсифицируется. 4. Градиент давления обмена диоксида углерода меньше, чем градиент давления обмена кисло рода, в то же время растворимость СО^ в легочной мембране в 20 раз больше, чем растворимость кислорода, поэтому он легче диффундирует через нее даже при меньшем градиенте давления. Насыщение гемоглобина Каждая молекула гемоглобина может связывать 4 молекулы кислорода, образуя оксигемоглобин; гемоглобин, который не связывается с кислородом, называется дезоксигемоглобином. Интенсивность соединения кислорода с гемоглобином зависит от Ро^ крови и силы связи между ними. На рис. 9.7, о приведена кривая диссоциации кислород-гемоглобина, демонстрирующая насыщенность гемоглобина при различных показателях ^*о,. Высокое Ро крови обеспечивает почти полное насыщение гемоглобина кислородом, т.е. максимальное количество присоединяемого гемоглобином кислорода. При понижении Р^ уменьшается и насыщенность гемоглобина. На насыщенность гемоглобина влияет множество факторов. Если, например, кровь становится более кислой, кривая диссоциации смещается вправо. Это показывает, что на тканевом уровне от гемоглобина отделилось большее количество кислорода. Смещение кривой вправо (рис. 9.7, 6) вследствие снижения рН называется эффектом Бора [б]. рН в легких, как правило, высокий, поэтому гемоглобин, проходящий по легким, может " рассчитывать" на значительное насыщение 180 Рис. 9.7. Обычная кривая диссоциации кислород-гемоглобина (а) и влияние р Н крови (б) и температуры крови (в) на ее форму: 1 — высокий рН крови (низкое Рсо^)', 2 — нормальный рН крови (нормальное рсо^);
3 — низкий рН крови (высокое Рсо^) кислородом. С другой стороны, на уровне тканей рН ниже, что приводит к диссоциации кислорода и гемоглобина и доставке кислорода к тканям. При физической нагрузке способность " выгружать" кислород в мышцы увеличивается, поскольку рН мышц снижается. Температура крови также влияет на диссоциацию кислорода. Как видно из рис. 9.7, в, повышение температуры крови вызывает смещение кривой диссоциации вправо, что свидетельствует о более интенсивной " отгрузке" кислорода. Вслед- 20 40 60 80 1(ю ствие этого гемоглобин отдает больше кислоро-а да, когда кровь проходит по активным мышцам, " согретым" вследствие метаболических процессов. В легких, где температура крови ниже, процесс насыщения гемоглобина кислородом более интенсивен.
Повышенная температура крови и концентрация ионов водорода (Н^ в работающих мышцах влияют на кривую диссоциации кислорода, обеспечивая его увеличенную доставку к активным мышцам
181 ко во время физической нагрузки (интенсивные усилия аэробного характера), вызывающей ограничение транспорта кислорода, пониженное содержание кислорода в крови уменьшает образование энергии и ограничивает мышечную деятельность. ТРАНСПОРТ ДИОКСИДА УГЛЕРОДА Транспорт диоксида углерода также осуществляется кровью. Выделенный из клеток углекислый газ транспортируется кровью главным образом в одной из трех форм: 1) растворенный в плазме; 2) в виде ионов бикарбоната, образовавшихся вследствие диссоциации угольной кислоты; 3) связанный с гемоглобином. Рассмотрим каждый вид транспорта. РАСТВОРЕННЫЙ ДИОКСИД УГЛЕРОДА Определенное количество диоксида углерода, выделенного из тканей, растворяется в плазме. Однако в таком виде транспортируется очень незначительное его количество (всего 7— 10 %). Растворенный диоксид углерода выделяется из плазмы в участке с пониженным ^со,, например, в легких. Там он диффундирует из капилляров в альвеолы и выводится из организма. Ионы бикарбоната Большая часть диоксида углерода (около 60 — 70 %) транспортируется в виде ионов бикарбоната. Молекулы диоксида углерода и воды, соединившись, образуют угольную кислоту (Н^СОд). Эта кислота нестабильна и быстро разлагается, высвобождая ион водорода (Н" 1" ), и образуя ион бикарбоната (НСОр СО, + Н^О -> Н^СОд -> Н-*- + НСО;. угольная ион кислота бикарбоната Н" *" затем связывается с гемоглобином, что вызывает эффект Бора, уже упоминавшийся выше, вследствие которого кривая диссоциации кислород-гемоглобина смещается вправо. Таким обра- Большая часть образованного активными мышцами диоксида углерода транспортируется обратно в легкие в виде ионов бикарбоната зом, образование иона бикарбоната способствует " разгрузке" кислорода. Посредством этого механизма гемоглобин действует как буфер, связывая и нейтрализуя Н" 1' и тем самым предотвращая значительное подкисление крови. Более подробно вопрос кислотно-щелочного равновесия рассматривается ниже. Когда кровь поступает в легкие, где ^со, ниже, Н'1" и ионы бикарбоната снова соединяются, образуя угольную кислоту, которая затем расщепляется на диоксид углерода и воду Н^СОд -> СО;, + Нр.
Снова образовавшийся таким образом диоксид углерода может попасть в альвеолы и выделиться из организма. Карбаминогемоглобин Транспорт диоксида углерода происходит также, когда газ связывается с гемоглобином, образуя соединение, которое называется карбаминогемог-лобин. Такое название обусловлено тем, что диоксид углерода связывается с аминокислотами в гло-биновой части молекулы гемоглобина, а не с ге-мовой группой, как кислород. Поскольку диоксид углерода соединяется с другой частью молекулы гемоглобина, чем кислород, конкуренции между обоими процессами не происходит. Связывание диоксида углерода зависит от оксигенации гемоглобина (дезоксигемоглобин значительно легче связывает диоксид углерода, чем оксигемоглобин) и парциального давления СО^ (диоксид углерода выделяется из гемоглобина при низком ^со; ). Таким образом, в легких, где ^" со, низкое, диоксид углерода легко выделяется из гемоглобина, попадает в альвеолы и выводится из организма. В ОБЗОРЕ... 1. Кислород транспортируется кровью, главным образом связанным с гемоглобином (в виде оксигемоглобина), хотя небольшое его количество транспортируется растворенным в плазме. 2. Насыщение гемоглобина кислородом снижается при • снижении -рсо,; • уменьшении рН; • повышении температуры. Каждое из этих условий может отражать повышенную локальную потребность в кислороде. Они обеспечивают " отгрузку" кислорода в нужный участок. 3. Насыщенность гемоглобина кислородом составляет, как правило, 98 %. Этот показатель отражает значительно большую концентрацию кислорода, чем требуется нашему организму, вследствие чего кислородтранспортная способность крови крайне редко ограничивает мышечную деятельность. 4. Диоксид углерода транспортируется кровью главным образом в виде ионов бикарбоната. Это предотвращает образование угольной кислоты, которая может вызвать аккумуляцию Н4', снижая Ш рН. Небольшое количество диоксида углерода транспортируется растворенным в плазме либо связанным с гемоглобином. ГАЗООБМЕН В МЫШЦАХ Итак, мы выяснили, как респираторная и сердечно-сосудистая системы обеспечивают доставку воздуха в легкие, обмен кислорода и диоксида углерода в альвеолах, транспорт кислорода к мышцам (и выведение из них диоксида углерода). Нам осталось рассмотреть, как транспортируется кислород из капиллярной крови к мышечным тканям и как выводится из них диоксид углерода. Этот газообмен между тканями и кровью в капиллярах представляет собой четвертый и последний этап транспорта газов — внутреннее дыхание. АРТЕРИОВЕНОЗНАЯ РАЗНИЦА ПО КИСЛОРОДУ В состоянии покоя содержание кислорода в артериальной крови составляет 20 мл/100 мл крови. Как видно из рис. 9.8, а, этот показатель снижается до 15—16 мл/100 мл крови, когда кровь проходит по капиллярам в венозную систему. Различие в содержании кислорода в артериальной и венозной крови называют артериовенозной разницей по кислороду (АВР—0^). Она отражает потребление тканями 4 — 5 мл кислорода/100 мл крови. Количество потребляемого кислорода прямо пропорционально количеству, используемому для образования энергии путем окисления. Следовательно, с увеличением использования кислорода АВР—0^ также возрастает. Например, при интенсивной физической нафузке, как видно из рис. 9.8Д АВР—0^ в сокращающихся мышцах может увеличиваться до 15 — 16 мл/100 мл крови. При ^ Артериовенозная разница по кислороду уве-у личивается с 4—5 мл/100 мл крови в состоянии покоя до 15—16 мл/100 мл крови при интенсивной физической нагрузке. Это увеличение отражает повышенное извлечение кислорода из артериальной крови активными мышцами, приводящее к снижению содержания кислорода в венозной крови таком усилии кровь отдает больше кислорода активным мышцам, поскольку в них намного Р.. ниже, чем в артериальной крови. ФАКТОРЫ, ВЛИЯЮЩИЕ НА ДОСТАВКУ И ИСПОЛЬЗОВАНИЕ КИСЛОРОДА Интенсивность доставки и использования кислорода зависит от трех переменных: 1) содержания кислорода в крови; 2) величины кровотока; 3) локальных усилий. Когда мы начинаем выполнять упражнение, каждая из этих трех переменных претерпевает изменения, направленные на обеспечение доставки большего количества кислорода к активным мышцам. Мы рассмотрели содержание кислорода в крови и знаем, что в обычных условиях насыщенность гемоглобина кислородом составляет 98 %. Любое снижение обычной кислородтранс-портной способности крови приведет к нарушению транспорта кислорода и уменьшению его клеточной утилизации. Как мы установили в главе 8, физическая нагрузка усиливает кровоток в мышцах. Поскольку большее количество крови транспортирует кислород через мышцы, из каждых 100 мл крови утилизируется меньшее количество кислорода (при условии, что потребность не изменилась). Следовательно, усиленный кровоток повышает эффективность доставки и потребления кислорода. Многие локальные изменения в мышце при выполнении физической нагрузки влияют на доставку и потребление кислорода. Например, мышечная активность повышает ее кислотность вследствие образования лактата. Кроме того, в результате усиленного метаболизма повышаются температура мышцы и концентрация диоксида углерода. Эти изменения увеличивают " отгрузку" кислорода из молекулы гемоглобина, способствуя его доставке и потреблению мышцами. При максимальной физической нагрузке, когда наш организм работает " на пределе" своих возможностей, изменение любой из этих переменных может нарушить транспорт кислорода и способность организма удовлетворять потребности окисления. Речь об этом пойдет дальше. 183
ВЫВЕДЕНИЕ ДИОКСИДА УГЛЕРОДА Диоксид углерода " покидает" клетки в результате диффузии в ответ на градиент парциального давления между тканью и кровью в капиллярах. К примеру, мышцы образуют диоксид углерода в результате окислительного метаболизма, следовательно, Р^ в мышцах будет относительно выше, чем в капиллярной крови. Поэтому СО^ диффундирует из мышц в кровь и транспортируется в легкие. Итак, мы завершили рассмотрение того, как кислород попадает в организм, транспортируется в ткани и как диоксид углерода выводится из тканей, транспортируется в легкие и выводится из организма. Кратко эти процессы показаны на рис. 9.9. 184 Кроме хеморецепторов и другие нервные механизмы влияют на дыхание. В плевре, бронхиолах и альвеолах находятся тензорецепторы. При чрезмерном растяжении этих участков соответствующая информация передается в центр выдоха. Он реагирует, сокращая продолжительность вдоха и тем самым снижая риск чрезмерного наполнения дыхательных структур. Это так называемый рефлекс Геринг—Брейера. Мы можем в некоторой степени произвольно контролировать дыхание с помощью мотонейронов коры головного мозга. Однако его может " перекрыть" непроизвольный контроль со стороны дыхательного центра. Попытайтесь задержать ды хание на 5 мин. В определенный момент, несмотря на ваше сознательное усилие задерживать дыхание, в организме достаточно повысятся уровни диоксида углерода и Н+, содержание кислорода понизится и центр вдоха заставит вас сделать вдох. Итак, как следует из рис. 9.10, в регуляции дыхания принимают участие многие механизмы. Оказывают влияние даже такие простые стимулы, как эмоциональный дистресс или резкое изменение температуры окружающей среды. Все эти механизмы необходимы. Цель дыхания — поддержание соответствующего количества газов в крови и тканях, а также соответствующего рН для обеспечения нормальной клеточной деятельности. Даже
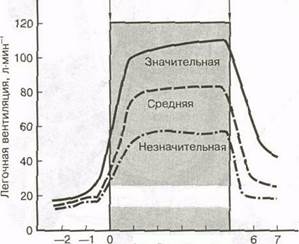
Рис. 9.10. Процессы, участвующие в регуляции дыхания 185 незначительные изменения этих переменных могут существенно повлиять на уровень мышечной деятельности и нанести вред здоровью. ЛЕГОЧНАЯ ВЕНТИЛЯЦИЯ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ Начало мышечной деятельности сопровождается усилением легочной вентиляции в два раза. Существенное увеличение происходит почти немедленно, затем следует продолжающееся постепенное увеличение глубины и частоты дыхания. Усиление вентиляции при выполнении физической нагрузки разной интенсивности иллюстрирует рис. 9.11. Подобное двухфазное увеличение свидетельствует о том, что первоначальное усиление вентиляции обусловлено механикой движений тела. С началом упражнения, прежде чем происходит любое химическое стимулирование, более активной становится двигательная область коры головного мозга, которая посылает стимулирующие импульсы в центр вдоха; он реагирует на них усилением дыхания. Кроме этого, механизм проприоцептивной обратной связи активных скелетных мышц и суставов обеспечивает дополнительную импульсацию, на которую также реагирует дыхательный центр. Вторая фаза увеличения дыхания обусловлена изменением температуры и химического состава артериальной крови. По мере выполнения физической нагрузки вследствие усиления метаболизма в мышцах образуется больше тепла, диоксида углерода и Н\ Все это способствует " разгрузке" мышц и повышает АВР—0^. Кроме того, в кровь попадает больше диоксида углерода, в результате чего увеличивается содержание в ней не только диоксида углерода, но и Н^ Это ощущают хемо- Начало Конец
12345 Время, мин Рис. 9.11. Вентиляторная реакция на значительную, среднюю и незначительную физическую нагрузку рецепторы, которые, в свою очередь, стимулируют дыхательный центр, повышая частоту и глубину дыхания. По мнению некоторых ученых, в этом процессе могут принимать участие и хемо-рецепторы мышц. Более того, по имеющимся данным рецепторы, находящиеся в правом желудочке, также посылают информацию в дыхательный центр, в результате чего увеличивается сердечный выброс, стимулирующий дыхание в первые минуты выполнения упражнения. Легочная вентиляция увеличивается при физической нагрузке, достигающей почти максимальной интенсивности, прямо пропорционально метаболическим потребностям организма. При более низкой интенсивности нагрузки это осуществляется за счет увеличения дыхательного объема — объема воздуха, вдыхаемого и выдыхаемого при нормальном дыхании. При увеличении интенсивности нагрузки частота дыхания также повышается. Максимальная величина легочной вентиляции зависит от размеров тела. У крупных людей максимальная вентиляция превышает 200 л-мин^, тогда как у людей с небольшими размерами тела она составляет около ]00л-мин~1 При прекращении физической нагрузки потребности мышц в энергии почти моментально снижаются до уровней, характерных для состояния покоя. В то же время легочная вентиляция возвращается к обычному уровню относительно медленнее. Если частота дыхания максимально соответствует метаболическим потребностям тканей, она снизится до исходного уровня в течение нескольких секунд после завершения физической нагрузки. Однако для восстановления дыхания потребуется несколько минут, что свидетельствует о том, что процесс дыхания после физической нагрузки регулируется главным образом кислотно-щелочным равновесием, ^сод и температурой крови. ПРОБЛЕМЫ ДЫХАНИЯ ПРИ ВЫПОЛНЕНИИ ФИЗИЧЕСКОЙ НАГРУЗКИ Самым идеальным вариантом регуляции дыхания при выполнении физической нагрузки был бы тот, который позволил бы оптимизировать нашу способность выполнять работу. К сожалению, это не всегда случается. При выполнении физической нагрузки могут возникать различные проблемы, связанные с дыханием, отрицательно влияющие на уровень мышечной деятельности. Рассмотрим некоторые из них. 186 Одышка Одышку при выполнении физической нагрузки очень часто испытывают физически плохо подготовленные люди, которые пытаются работать с интенсивностью, приводящей к значительному повышению уровня артериального диоксида углерода и концентрации Н+. Как уже указывалось, оба эти стимула посылают сигналы в дыхательный центр увеличить частоту и глубину вентиляции. Хотя одышка, обусловленная физической нагрузкой, представляет собой затрудненное дыхание, она вызвана неспособностью адаптации ^со, крови и Н+. Неспособность снизить действие этих стимулов во время физической нагрузки, по-видимому, связана с недостаточной подготовленностью дыхательных мышц. Несмотря на значительное побуждение к вентиляции легких, дыхательные мышцы легко устают и не могут восстановить нормальный гомеостаз. кислорода в центральную нервную систему. Преимущества гипервентиляции перед физической нагрузкой не совсем выяснены, гипервентиляция может скорее отрицательно повлиять на физическую деятельность, чем улучшить ее. Возможно, ответ на этот вопрос дадут результаты будущих исследований. Гипервентиляцию практикуют также ныряльщики. Гипервентиляция снижает побуждение сделать вдох, однако не увеличивает запасы кислорода в организме. Дальнейшая задержка дыхания становится невозможной, когда /^со, в артериальной крови достигает 55 мм рт.ст. К сожалению, при погружении под воду, которому предшествовала гипервентиляция, содержание кислорода в крови может снизиться до критических уровней задолго до того, как аккумуляция СО^ " даст сигнал" подниматься на поверхность и сделать вдох. Человек может потерять сознание, прежде чем у него возникнет потребность сделать вдох. Гипервентиляция Некоторые респираторные расстройства, а также предстартовое волнение могут вызвать резкое увеличение вентиляции, превышающее метаболические потребности в кислороде. Это явление называется гипервентиляцией. В состоянии покоя произвольная гипервентиляция приводит к снижению нормального показателя Рсо^, — 40 мм рт.ст. — в альвеолах и артериальной крови до 15 мм рт.ст. Поскольку снижается уровень диоксида углерода, то уменьшается и количество Н+, в результате чего увеличивается рН крови. Эти изменения снижают вентиляторное побуждение. Поскольку кровь, идущая из легких, почти всегда на 98 % насыщена кислородом, увеличение альвеолярного ^со, не повышает содержание кислорода в крови. Поэтому пониженное стремление дышать в сочетании с повышенной способностью задержать дыхание после гипервентилирования —результат " разгрузки" диоксида углерода, а не повышенного содержания кислорода в крови. Глубокое быстрое дыхание в течение нескольких секунд может привести к головокружению и даже потере сознания. Это явление демонстрирует чувствительность регуляции респираторной системой содержания диоксида углерода и рН. В надежде снизить респираторный дистресс-синдром пловцы довольно часто перед соревнованием осуществляют гипервентиляцию. Задержка дыхания во время плавания благоприятно влияет на механику гребка, поэтому многие пловцы-спринтеры проводят гипервентиляцию непосредственно перед стартом. Хотя в первые 8— 10 с заплыва у них практически не возникает желание сделать вдох, содержание кислорода в альвеолах и артериях может понизиться до критического, так как кислород используется, но не пополняется. Это может нарушить мышечное окисление и транспорт Проба Вальсальвы Дыхательная процедура, которую часто применяют при выполнении определенных физических упражнений и которая может быть весьма опасной, называется пробой Вальсальвы. Она включает следующие этапы: 1) перекрытие голосовой щели; 2) увеличение внутрибрюшного давления вследствие мощного сокращения диафрагмы и мышц живота; 3) увеличение внутри грудного давления за счет мощного сокращения дыхательных мышц. В результате этих действий воздух задерживается в легких и оказывается под высоким давлением. Пробу Вальсальвы часто используют при поднятии тяжелых объектов, когда человек пытается стабилизировать грудную клетку. Высокое внутрибрюшное и внутригрудное давление ограничивает венозный возврат, коллапси-руя большие вены. Если проба Вальсальвы продолжается длительное время, объем крови, возвращающейся в сердце, значительно уменьшается, приводя к снижению сердечного выброса. Хотя в некоторых случаях этот опыт дает положительные результаты, он может оказать и весьма серьезное отрицательное воздействие. Людям, страдающим гипертензией или другими сердечно-сосудистыми расстройствами, не следует практиковать пробу Вальсальвы. В ОБЗОРЕ... 1. Дыхательные центры, расположенные в стволе головного мозга, задают частоту и глубину дыхания. 2. Центральные хеморецепторы головного моз- 187
га реагируют на изменения концентраций диоксида углерода и Н+. При повышении любой из этих переменных центр вдоха усиливает дыхание. 3. Периферические рецепторы, расположенные на дуге аорты и разветвлении сонной артерии, реагируют главным образом на изменение содержания кислорода, СО^ и Н^ При значительном снижении содержания кислорода или повышении уровней Н4' и СО^ они передают эту информацию центру вдоха, который усиливает дыхание. 4. Тензорецспторы дыхательных путей и легких вынуждают центр выдоха сократить дыхание, чтобы не допустить избыточного наполнения легких. Кроме того, человек в определенной степени может произвольно контролировать дыхание. 5. Во время физической нагрузки вентиляция усиливается почти сразу же в результате стимуляции центра вдоха, обусловленной самой мышечной деятельностью. После этого следует более постепенное ее увеличение вследствие повышения температуры и химических изменений в артериальной крови в результате мышечной деятельности. 6. Проблемы дыхания, возникающие при выполнении физической нагрузки, включают одышку, гипервентиляцию и осуществление пробы Вальсальвы. ВЕНТИЛЯЦИЯ И ОБМЕН ЭНЕРГИИ При продолжительном выполнении мышечной деятельности средней интенсивности вентиляция, как правило, соответствует интенсивности обмена энергии. Она имеет тенденцию изменяться прямо пропорционально объему потребляемого кислорода и диоксида углерода, производимого организмом. Рассмотрим, насколько тесно дыхание связано с потреблением кислорода. ВЕНТИЛЯЦИОННЫЙ ЭКВИВАЛЕНТ ПО КИСЛОРОДУ Отношение объема вентилируемого воздуха (У^) к количеству поглощаемого тканями кислорода (Уо^) иллюстрирует экономичность дыхания. Это отношение называют вентиляционным эквивалентом по кислороду или у^ /у() • Он измеряется количеством воздуха, которым мы дышим (в литрах), к количеству потребляемого кислорода (в литрах). В состоянии покоя У^/Уо колеблется от 23 до 28 л воздуха на литр потребляемого кислорода. Этот показатель незначительно изменяется при умеренной нагрузке, например, ходьбе. Однако с увеличением интенсивности нагрузки до почти максимальных уровней У^/Уц может превышать 30 л воздуха/л кислорода. В принципе У^/Уа остается относительно постоянным при значитель ном колебании уровней нагрузки. Это свидетельствует о том, что системы регуляции дыхания адекватно реагируют на потребности организма в кислороде. Даже в таких видах спорта, как плавание, где дыхание должно осуществляться синхронно с циклом гребка рукой, У^/Уо, практически не отличается от показателей, характерных для других видов деятельности. МОМЕНТ СНИЖЕНИЯ ЭФФЕКТИВНОСТИ ВЕНТИЛЯЦИИ При повышении интенсивности физической нагрузки до максимальной в определенный момент вентиляция начинает увеличиваться непропорционально потреблению кислорода. Это так называемый момент снижения эффективности вентиляции (рис. 9.12). Когда интенсивность нагрузки превышает 55 — 70 % МПК, транспортируемый в мышцы кислород не удовлетворяет потребность в нем для окисления. Это компенсируется образованием большего количества энергии путем гликолиза, что ведет к увеличению образования и аккумуляции молочной кислоты. Молочная кислота в сочетании с бикарбонатом натрия, нейтрализующим кислоту, образуют лактат натрия, воду и диоксид углерода. Как нам уже известно, диоксид углерода стимулирует хеморецеп-торы, которые посылают сигнал в центр вдоха увеличить вентиляцию. Таким образом, момент снижения эффективности вентиляции отражает реакцию респираторной системы на повышенное содержание СО,. Как видно из табл. 9.2, после наступления этого момента вентиляция значительно повышается. 160
160 180 200 220 240 260 Скорость бега, м-мин" ' Рис. 9.12. Изменения легочной вентиляции во время выполнения физической нагрузки, иллюстрирующие момент снижения эффективности вентиляции: 1 — потребление кислорода; 2— вентиляция; 3— момент снижения эффективности вентиляции 188 Таблица 9.2. Соотношение между легочной вентиляцией V,. и потреблением кислорода Уц
Вентиляция увеличивается прямо пропорционально интенсивности выполнения работы до момента снижения ее эффективности. После этого она увеличивается непропорционально по мере того, как организм пытается избавиться от излишнего СО-
150 200 250 300 Скорость бега, м.мин" ' Рис. 9.13. Изменение вентиляторного эквивалента по диоксиду углерода (Уу/У^ ) и кислороду (Vр/Рд ) при повышении интенсивности бега АНАЭРОБНЫЙ ПОРОГ Одно время бытовало мнение, что ввиду непропорционального увеличения вентиляции без усиления потребления кислорода момент снижения эффективности вентиляции может быть связан с порогом лактата (начало аккумуляции лакта-та, превышающей уровни, характерные для состояния покоя, при выполнении физической нагрузки с постепенным увеличением интенсивности). Момент снижения эффективности вентиляции отражает увеличение объема диоксида углерода, производимого за 1 мин (Усо^). Вспомним из главы 5, что коэффициент дыхательного газообмена (КДГ) представляет собой соотношение образования диоксида углерода к потреблению кислорода. Следовательно, увеличение образования диоксида углерода приводит к повышению КДГ. Считалось, что повышенный ^со, — результат чрезмерного выделения диоксида углерода из бикарбоната, нейтрализующего молочную кислоту. Вассерман и Мак-Илрой ввели понятие анаэробного порога для характеристики этого явления, поскольку считали, что резкое увеличение количества СО^ отражает смещение в сторону более анаэробного метаболизма [10]. Они использовали увеличение КДГ в качестве маркера анаэробного порога, считая его достаточной альтернативой пробе крови, позволяющей определить начало анаэробного метаболизма. За многие годы этот подход постоянно совершенствовался. В настоящее время наиболее точный метод идентификации анаэробного порога основывается на показателях вентиляторного эквивалента по кислороду (У^/Уц ) и диоксиду уг лерода (У^/Усо )• Последний представляет собой отношение объема вдыхаемого воздуха к объему образуемого СО^. Наиболее специфичный критерий оценки анаэробного порога — системное увеличение У^/Уц без сопутствующего повышения ^Е/^со [2] (рис. 9.13). Вентиляторный эквивалент по диоксиду углерода относительно постоянен, что свидетельствует о соответствии вентиляции потребности организма в выведении СОд. Увеличение У^/Уо свидетельствует, что усиление вентиляции, направленное на выведение СО,, непропорционально потребности организма в Од. ™У Анаэробный порог в большинстве случаев • достаточно точно отражает порог лактата, вместе с тем взаимосвязь их недостаточно совершенна Анаэробный порог служит неинвазивной оценкой порога лактата и в большинстве случаев они оба наблюдаются в один и тот же момент времени при выполнении физической нагрузки с увеличивающейся интенсивностью или при одном и том же МПК. Однако есть и исключения [I]. Например, у людей, страдающих болезнью Мак-Ар-дла, уровни лактата крови и Н'1' не повышаются во время физической нагрузки вследствие отсутствия мышечной фосфорилазы. У них отмечается четкий анаэробный порог при выполнении физической нагрузки с увеличивающейся интенсивностью, несмотря на то, что концентрация лактата крови находится на уровне, характерном для 189 состояния покоя. Истощение запасов гликогена перед физической нагрузкой также изменяет взаимосвязь анаэробного порога и порога лактата. В ОБЗОРЕ... 1. При выполнении физической нагрузки средней степени интенсивности вентиляция достаточно точно отражает интенсивность обмена энергии и соответствует потреблению кислорода. Отношение объема вентилируемого воздуха к потреблению кислорода представляет собой вентиляторный эквивалент по кислороду (У^/Уц )• 2. Момент снижения эффективности вентиляции — резкое усиление вентиляции без соответствующего увеличения потребления кислорода. Он отражает потребность выведения избыточного количества диоксида углерода. 3. Анаэробный порог соответствует моменту, когда У^/У» резко повышается, тогда как У^/Уса, остается относительно стабильным. Анаэробный порог представляет собой неинвазивный метод определения порога лактата. 190 ют отрицательное воздействие на людей, страдающих астмой. Несмотря на интенсивные исследования, механизм или механизмы, вследствие которых физические нагрузки могут приводить к нарушению проходимости дыхательных путей у людей, страдающих астмой, до сих пор не установлены. Более подробно эта проблема освещается в работах Слая и Эгглстона [4, 9]. В ОБЗОРЕ... 1. При значительной физической нагрузке дыхательные мышцы могут использовать более 15 % кислорода, потребляемого организмом. 2. Легочная вентиляция обычно не является фактором, лимитирующим мышечную деятельность даже при максимальном усилии. Об этом может идти речь лишь в отдельных случаях у отлично тренированных спортсменов. 3. По-видимому, дыхательные мышцы лучше приспособлены " избегать" утомления при продолжительной физической работе, чем мышцы конечностей. 4. Сопротивление дыхательных путей и диффузия газов, как правило, не являются ограничительными факторами мышечной деятельности у физически здоровых людей. 5. Респираторная система может ограничивать мышечную деятельность у людей с респираторными нарушениями — нарушением проходимости или сужением дыхательных путей. РЕСПИРАТОРНАЯ РЕГУЛЯЦИЯ КИСЛОТНО-ЩЕЛОЧНОГО РАВНОВЕСИЯ Как уже отмечалось, интенсивная мышечная деятельность довольно часто приводит к образованию и аккумуляции лактата и Н" 1". Это может отрицательно повлиять на обмен энергии, а также снизить силу мышечных сокращений. Хотя регуляция кислотно-щелочного равновесия в организме осуществляется не только благодаря респираторной системе, мы решили включить ее рассмотрение в этот раздел, поскольку она играет очень важную роль в быстрой адаптации кислотно-щелочного статуса во время и сразу же после мышечной деятельности. Кислоты, такие, как молочная и угольная, выделяют ионы водорода (Н4'). Как отмечалось в главе 5, вследствие метаболизма углеводов, жиров или белков образуются неорганические кислоты, которые, расщепляясь, повышают концентрацию Н+ в жидкостях организма. Для нейтрализации действия свободных Н'1' в крови и мышцах содержатся основные вещества, которые, вступая в реакцию, нейтрализуют Н" ^: Н+ + буфер -> Н-буфер. В состоянии покоя в жидкостях организма этих веществ (таких, как бикарбонат, фосфат и белки) содержится больше, чем кислот. В результате рН колеблется от 7, 1 в мышцах до 7, 4 в артериальной крови. Переносимые (в течение всего нескольких минут) пределы рН артериальной крови составляют 6, 9 — 7, 5 (рис. 9.14). Превышающая нормальную концентрация Н4" называется ацидозом, тогда как концентрация ниже нормальной — алкалозом. рН внутри- и внеклеточных жидкостей организма находится в относительно узком диапазоне благодаря химическим буферам, легочной вентиляции и функции почек. Три основные химические буфера в организме — бикарбонат (НСО^), фосфаты (Р^) и белки. Кроме них, как отмечалось ранее, одним из главных буферов является гемоглобин эритроцитов. Табл. 9.3 иллюстрирует относительный вклад этих буферов в регуляцию концентрации кислот в крови. Вспомним, что бикарбонат, соединяясь с Н_^, образует угольную кислоту, тем самым снижая ее подкисляющее влияние. Угольная кислота, в свою очередь, образует диоксид углерода и воду в легких. СО^ затем выводится, а вода остается. Таблица 9.3. Буферная способность компонентов крови Буфер Слайксы* Бикарбонат
Гемоглобин Белки Фосфаты Итого 28, 0 *Миллиэквиваленты ионов водорода, потребляемые каждым литром при рН 7, 4 — 7, 0.
б Рис. 9.14. Допустимые пределы рН артериальной крови (а) и мышц (б) в покое и в состоянии изнеможения. Обратите внимание на ограниченный диапазон физиологической толерантности для рН крови и мышц: 1 — допустимые пределы для артериальной крови (6, 9 — 7, 5); 2 — состояние изнеможения (6, 63); 3 — состояние покоя (7, 10) 191 Количество бикарбоната, соединяющегося с Н" '", равно количеству буферируемой кислоты. Когда молочная кислота приводит к снижению рН от 7, 4 до 7, 0, используется более 60 % бикарбоната, первоначально содержавшегося в крови. Даже в состоянии покоя кислота, образуемая конечными продуктами обмена веществ, извлекает из крови большую часть бикарбоната, если нет другого способа выведения Н4 из организма. К счастью, кровь и буферы требуются только для транспорта кислот, образуемых в результате обмена веществ, из участков их образования (мышц) в легкие или почки, откуда они выводятся. После завершения транспорта кислот буферные молекулы могут быть снова использованы. В мышечных волокнах и почечных канальцах Н" ^ в основном буферируется фосфатами — фосфорной кислотой и фосфатом натрия. В настоящее время мало сведений о способностях буферов клеток. Нам только известно, что в клетках содержится больше белков и фосфатов и меньше бикарбоната, чем во внеклеточной жидкости. Как уже отмечалось, любое увеличение количества свободного Н'1' в крови стимулирует дыхательный центр усилить вентиляцию. Это способствует связыванию Н4' и бикарбоната и выведению СО-. Конечным результатом является снижение содержания свободных Н'1' и увеличение рН крови. Таким образом, как химические буферы, так и респираторная система обеспечивают Загрязнение воздуха В последние 20 лет озабоченность специалистов вызывает проблема, связанная с осуществлением мышечной деятельности в условиях загрязненного воздуха. Воздух во многих городах загрязнен небольшим количеством газов и частиц, которые не должны содержаться в воздухе, которым мы дышим. Когда воздух становится спертым или происходит инверсия температуры, концентрация некоторых из этих загрязняющих веществ повышается, что в значительной мере влияет на спортивную деятельность. Основными загрязняющими веществами в данном случае являются оксид углерода, озон и оксид серы. Оксид углерода (СО) — газ, не имеющий запаха, который при вдыхании быстро попадает в кровь и может привести к летальному исходу. Сродство гемоглобина с оксидом углерода почти в 240 раз выше, чем с кислородом, поэтому гемоглобин " предпочитает" связать как раз оксид углерода. Уровни оксида углерода в крови непосредственно определяются его содержанием во вдыхаемом воздухе. В ряде исследований отмечали линейное снижение МПК при увеличении содержания оксида углерода в крови. На основании анализа литературы по данному вопросу можно сделать вывод, что снижение МПК не является статистически значимым до тех пор, пока количество оксида углерода в крови не превысит 4, 3 %; вместе с тем результаты теста на тредбане понижаются при уровне оксида углерода 2, 7 % [5, 8]. Содержание оксида углерода в крови свыше 15 % влияет на выполнение субмаксимальной нагрузки при интенсивности менее 60 % МПК. Озон (0^) — наиболее типичный фотохимический окислитель. При высокой концен трации озона во вдыхаемом воздухе могут возникать кашель, раздражение глаз, одышка, рвота и т.д. Озон, в первую очередь, воздействует на дыхательные пути. Нарушение функции легких наблюдается при повышении его концентрации в окружающем воздухе. МПК значительно снижается при интервальном выполнении физической нагрузки в течение 2 ч при воздействии озона порядка 0, 75 %о. Это снижение МПК, очевидно, связано с пониженным транспортом кислорода в легкие вследствие менее интенсивного альвеолярного газообмена. Еще одно загрязняющее вещество — оксид серы (80^). О нем известно немного. Если концентрация данного газа в воздухе превышает 1, 0 %о, это приводит к значительному дискомфорту и существенному снижению спортивных результатов [8]. Оксид серы раздражает в основном верхние дыхательные пути и бронхи. В некоторых городах начали проводить кампанию, направленную на предупреждение жителей о степени загрязненности воздуха и смоге. Она предполагает использование специального цветного кодирования, где каждый цвет соответствует степени загрязнения воздуха. В этой связи целесообразно разработать единые стандарты для контроля за чистотой воздуха. Имеющиеся данные убедительно показывают необходимость отмены всех спортивных состязаний, когда уровень загрязнения воздуха превышает безопасные пределы. Мы надеемся, что последующие исследования позволят лучше понять отрицательные воздействия загрязняющих веществ на мышечную деятельность. 192 временную нейтрализацию срочных воздействий ацидоза вследствие физической нагрузки. Для поддержания постоянного буферного резерва накопившиеся Н^ выводятся из организма через почки и мочевую систему. Почки фильтруют Н+ из крови вместе с другими продуктами распада. Это обеспечивает выведение Н'1' из организма при сохранении концентрации внеклеточного бикарбоната. При кратковременной физической нагрузке (спринт) в мышцах образуется большое количество лактата и Н+, которые снижают рН мышц с 7, 08, характерного для состояния покоя, до 6, 70. Как видно из табл. 9.4, бег с максимальным усилием на дистанцию 400 м приводит к снижению рН мышц ног до 6, 63 и повышению концентрации лактата с 1, 2 до 19, 7 ммоль-кг" ' мышцы. Как уже отмечалось, подобное нарушение кислотно-щелочного равновесия отрицательно влияет на сократительные способности мышц и их способность образовывать АТФ. Накопление лактата и Н" 1" частично обусловлено тем, что они не могут легко диффундировать через мембраны волокон. Несмотря на значительное образование лактата и Н^ в течение 60 с, необходимых, чтобы пробежать дистанцию 400 м, эти промежуточные продукты диффундируют через жидкости организма и достигают равновесия только через 5—10 мин во время восстановления. Через 5 мин после забега у бегунов (табл. 9.4) показатели рН крови были 7, 10, а показатели лактата— 12, 3 ммоль-л" ', по сравнению с 7, 40 и 1, 5 ммоль-л~1 в состоянии покоя. Таблица 9.4. рН и концентрация лактата в крови и мышцах после забега на 400 м
20 40 60 80 Восстановление, мин Рис. 9.15. Влияние активного и пассивного восстановления уровней лактата крови после серии спринтерских забегов: 1 — пассивное восстановление; 2 — активное восстановление; 3 — уровень в состоянии покоя ток в активных мышцах, который, в свою очередь, усиливает диффузию лактата мышц и его окисление. Хотя уровень лактата крови остается повышенным в течение 1 — 2 ч после значительной физической нагрузки анаэробной направленности, концентрация Н'*" в крови и мышцах восстанавливается в течение 30— 40 мин. Это обусловлено химической нейтрализацией в основном бикарбоната и респираторным выведением избытка СОу
Восстановление нормальных уровней лактата крови и мышц после такой изнурительной нагрузки — относительно длительный процесс, продолжающийся, как правило, 1 — 2 ч. Как видно из рис. 9.15, восстановлению уровня лактата крови способствует непрерывное выполнение физической нагрузки небольшой интенсивности, так называемое активное восстановление [7]. После серии спринтерских забегов испытуемые либо сидели неподвижно (пассивное восстановление), либо работали с интенсивностью 50 % МП К. Во время активного восстановления лактат крови также восстанавливается быстрее, поскольку мышечная деятельность поддерживает высокий крово- В ОБЗОРЕ... 1. Чрезмерная концентрация Н+ (пониженное рН) нарушает образование АТФ и сократительную способность мышц. 2. Респираторная система играет существенную роль в поддержании кислотно-щелочного равновесия. 3. При увеличении количества Н" *' центр вдоха реагирует усилением дыхания. Главный способ снижения концентрации Н'1' — выведение СОу Диоксид углерода транспортируется в основном в связанном с бикарбонатом виде. Достигнув легких, он диссоциирует и выводится из организма. 4. При повышении концентрации Н+ вследствие аккумуляции либо СО-, либо лактата ионы бикарбоната нейтрализуют Н+, предотвращая ацидоз. В главе 8 мы выяснили, какую роль играет сердечно-сосудистая система при выполнении физических нагрузок. В данной главе мы рассмотрели
роль респираторной системы. Выяснили, каким образом эта система может ограничивать мышечную деятельность. В следующей главе остановимся на адаптационных реакциях сердечно-сосудистой и респираторной систем вследствие физических нагрузок. Рассмотрим, как эти адаптационные реакции могут повышать способности обеих систем удовлетворять потребности организма и как они влияют на спортивную деятельность. Контрольные вопросы 1. Опишите анатомические структуры, участвующие в процессе легочной вентиляции. 2. Какие мышцы связаны с процессом дыхания, расскажите об их функциях во время легочной вентиляции. 3. Что такое парциальное давление кислорода и диоксида углерода во вдыхаемом воздухе, альвеолярном воздухе, артериальной и смешанной венозной крови? 4. В каком виде транспортируется кровью кислород и диоксид углерода? 5. Какие химические стимулы регулируют частоту и глубину дыхания? Как они контролируют дыхание во время физической нагрузки? Какое действие оказывает на них произвольная гипервентиляция? 6. Какие еще факторы регулируют вентиляцию во время физической нагрузки? 7. Что такое вентиляторный эквивалент по кислороду? по СО,? 8. Определите момент снижения эффективности вентиляции и анаэробный порог. 9. Что такое порог лактата, анаэробный порог? Как связаны эти два понятия? 10. Какую роль играет респираторная система в кислотно-щелочном равновесии? Я. Какой обычный (в состоянии покоя) показатель рН артериальной крови? Мышц? Как изменяется этот показатель при выполнении физической нагрузки спринтерской направленности? 12. Какие основные буферы крови и мышц вы знаете? 13. Какая продолжительность восстановления уровней рН и лактата крови после мышечной деятельности спринтерской направленности? СПИСОК ЛИТЕРАТУРЫ 1. Апйегеоп 0.8., КЬойез Е.С. (1989). А геу; е^ оГ Ыоо< 11ас1а1е апс1 уепН1а1огу тецюсЬ оГс1е1есНп§ (гапвШоп 1Ьге$Ьо1(15. §рой5 МеШсте, 8, 43 — 55. 2. Оау; & .1.А. (1985). АпаегоЫс Шге5Ьо1с1: Кеу1е\у оГ Ше сопсер! апс1 сНгейюпх Гог ГиШге ге$еагсп. МесИсте апс1 5с1епсе ш 5роП5 апс1 Ехегс1& е, 17, 6 — 18. 3. Оетр«, еу.1.А., Ук1ги1( Е.Н., МИспеП 0.5. (1986). Ь» 1Ье 1ипв Ьш11 Гое ехегс^е? МесИсте апс1 5с1епсе т 5рог1 апс1 Ехегс^е, 18, 143- 155. 4. Е§е1ех1оп Р.А. (1986). Ра(порпу5ю1о§у оГ ехегс^е-шйисей аЯтпа. МесКсте апс1 §с; епсе т 5роП апс1 Ехетзе, 18, 318- 321. 5. РоИпзЬее Ы., Кауеп Р.В. (1984). Ехегс; 5е апс1 аи-роИиНоп. }оита1 оГ ЗроПв 5с1епсе5, 2 57 — 75. 6. Оиу1оп А.С. (1991). Тех1Ьоо1< оГтеШса! рЬу5ю1о§у (8111 ей.). РЬНа(Зе1рЫа: Заипйегк. 7. Неппапкеп Ь. (1981). Е{Гес1оГте1аЬо1к: сЬапвевоп Гогсе §епегаПоп т 51< е1е1а1 ти5с1е Диппе тах1та1 ехеплве. 1п К.РоПег апс1 ^.\V^1е1ап (ЕДз.). Нитап ти5с1е ГаИеие: РЬу5; о1ов; са1 тесЬатктз (рр. 75 — 88). Ьоп(1оп: РИтап Ме(11са1. 8. Кауеп Р.В. (1979). Неа1 апс! а; г ро11и1юп: ТЬе сагШас раиет. 1п М.Ь. Ро11ос1< О.Н.§сЬтИ1 (ЕЙ5.), НеаП(115еахе апс1 гепаЫН1а1юп. Во51оп: НоиеЬюп М1гП1п. 9. 81у К.М. (1986). Н15(огу оГ ехегс; 5е-; пс1исе(1 ав1Ьта. Мес1ю1пе апс1 §с1епсе т ЗроПз апс1 Ехегс^е, 18, 314 — 317. 10. •\Уа55егтап К., МсПгоу М.В. (1964). Ое1ес1; п§ 1Ье 1Ьге5Ьо1с1 оГ апаегоЫс те1аЬоИ5т т сагсИас ра11еп(5 с1ипп§ ехегске. Атепсап ^оита1 оГ Сагс1ю1о§у, 14, 844— 852. РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА Вгоо! » О.А. (1985). Кеаропве ю Оау; 5 тапивспр!. МесИсте апс1 §с1епсе 1п ЗроПв апс1 Ехегс^е, 17, 19— 21. ВгооЬв С.А., Ра1Ьеу Т.О. (1985). Ехегс^е рЬу5ю1о§у: Нитап Ыоепег§е11с5 апс1 11& арр11са1юп& (рр. 221 — 278). N0^ УоА: \У11еу. Сотгое.1.Н. (1974). РЬуяо1о§у оГ ге5р{га1юп (2п< Э е< 1.). СЫсаео: Уеаг Воо1< МесИса!. Со^иИ О.Ь. (1970). Ме1аЬоЬс гезропвев с1иппв с1; 51ап& е гипп1п§. 1оита1 оГАррПес! РЬу5ю1оеу, 28, 251 — 255. Со5(Ш О.Ь., ВатеП А., §Ьагр К., Гт\и V/.!., Ка1г А. (1983). Еее тизс1е рН Го11о\уте 5рпп1 шпп1пв. Мес11с1пе апс1 §с1епсе 1п §рог15 апс1 Ехегс^е, 15, 325 — 329. СозШ! О.Ь., Уеге(арреп Р., Ки1рег5 Н., 1ап55еп Е., Р1п1( \У. (1984). АсИ-Ьаве Ьа1апсе с1ипп§ гереа(ес1 Ьои15 оГ ехегс^е: 1п(1иепсе оГ НСО. 1п1егпа1юпа1 ^оита1 оГ 8роП8 Мес1; с; пе, 5, 228 — 231. Оетрвеу ].А., Ук1ги1с Е.Н., Ма51епЬгоо1( 5.М. (1980). Ри1топагу соп1го1 5у51ет5 т ехегс1& е. Рес1ега1юп РгосеесИпев, 39, 1498 — 1505. Оетрзеу ^.А., У1(1ги1( Е.Н., МИсЬеП 0.5. (1985). Ри1топагу соп1го1 5у51ет2 1п ехегс1& е. Рес1ега1юп РгосеесИпез, 44, 2260 — 2270. Оетрвеу.1.0. (1985). Ехегс15е апй сЬетогесер11оп. Атепсап Кеу1е\у оГ Ке5р1га1огу 015еа5е, 129, 31 — 34. Ро\уеге 8.К., Но\у1еу Е.Т. (1990). Ехегс^е рЬу81о1оеу: Тпеогу апй арр11са1юп 1о П1пе85 апс1 регГогтапсе. ОиЬиоие, 1А: вго\уп. 8Ьаф К.Ь., СояпП О.Ь., Рт1( ^V.^., К1пе 0.8. (1986). Е(Гес15 оГе1§Ь1 \уее1< 5 оГЫсус1е егаотпег 5рпп11га1п1п§ оп Ьитап ти5с1е ЬиДег сарас11у. 1п1егпа1юпа1.1оигпа1 оГ8роп5 МесИсте, 7, 13— 17. 8и1Юп ^.К., ^опе& N.^., Тоеуух С.:. (1981). Епес1 оГрН оп ти8с1е §1усо1у5; 5 с1ипп§ ехегс15е. СИтса! 5с1епсе, 61, 331 - 338. ЛЕГОЧНАЯ ВЕНТИЛЯЦИЯ Легочная вентиляция, или просто дыхание, представляет собой процесс перемещения воздуха в легкие и из легких. Анатомическая структура респираторной системы показана на рис. 9.1. Воздух, как правило, поступает в легкие через нос; рот используется в том случае, когда потребность в воздухе превышает количество, которое может попасть в легкие через нос. Дыхание через нос предпочтительнее, поскольку воздух согревается и увлажняется, проходя через пространство неправильной формы (носовую раковину). Кроме того, при прохождении через носовую раковину пыль и другие частицы оседают на слизистой оболочке носоглотки. Таким образом осуществляется фильтрация всех частиц, за исключением мельчайших, что снижает раздражение и вероятность респираторных инфекций. Из носа и рта воздух движется по глотке, гортани, трахее, бронхам и бронхиолам, пока не 174
достигнет самых маленьких респираторных единиц — альвеол. Альвеолы являются местом газообмена в легких. Дыхание через нос обеспечивает согревание и увлажнение воздуха, а также фильтрацию инородных частиц Легкие не прикреплены непосредственно к ребрам, а " подвешены" с помощью плевральных полостей, окутывающих легкие. Они содержат тонкий слой плевральной жидкости, которая снижает трение при дыхательных движениях. Кроме
Рис. 9.1. Анатомия дыхательной системы того, эти полости соединены с легкими, а также с внутренней поверхностью грудной клетки, которая придает легким свой размер и форму по мере ее расширения и сокращения. Взаимосвязь между легкими, плевральными полостями и грудной клеткой определяет потоки вдыхаемого и выдыхаемого воздуха. Рассмотрим обе фазы —вдох и выдох. ВДОХ Вдох — активный процесс, в котором участвуют диафрагма и внешние межреберные мышцы. Динамика выполнения вдоха показана на рис. 9.2. Движение ребер и грудины осуществляется внешними межреберными мышцами. Ребра движутся вверх и в стороны подобно ручке ведра. Движения грудины направлены вверх и вперед и напоминают движения ручки насоса. Что касается диафрагмы, то она сокращается, опускаясь вниз к брюшной полости. Эти действия вызывают увеличение объема грудной клетки и легких. При расширении легких, воздух, находящийся в них, заполняет больше пространства и давление в легких снижается (рис. 9.2, в). В результате давление в легких (внутрилегочное давление) становится меньшим, чем давление окружающего воздуха. Поскольку дыхательные пути открыты, воздух устремляется в легкие, чтобы снизить разность давления. Таким образом при вдохе в легкие попадает воздух. В условиях выполнения значительной физической нагрузки осуществлению вдоха способствуют другие мышцы: лестничные (передняя, средняя и задняя) и грудино-ключично-сосцевидная, расположенные в области шеи, а также грудные. С их помощью ребра поднимаются выше, чем при обычном дыхании. Изменение давления, обеспечивающее адекватное вентилирование в состоянии покоя, очень незначительно. Например, при обычном атмосферном давлении (760мм рт.ст.) в результате вдоха давление в легких (внутрилегочное давление) может снизиться всего на 3 мм рт.ст. Однако при максимальном респираторном усилии, например, во время выполнения значительной физической нагрузки, внутрилегочное давление может снизиться на 80— 100 мм рт.ст.! ВЫДОХ В состоянии покоя выдох, как правило, — пассивный процесс, который включает расслабление дыхательных мышц и эластическую тягу легочной ткани (рис. 9.3, а). При расслаблении диафрагмы она принимает свое обычное дугообразное 175 Во время вдоха
Ребра движутся подобно Грудина движется вперед и ручке ведра, расширяя объемы груди вверх подобно ручке насоса, расширяя объемы груди (по бокам (спереди и сзади) (вид сбоку )
)
Мышцы сокращаются, легкие Вдох: воздух устремляется внутрь, уравновешивая давление расширяются: Давление в легких< атмосферного давление
Рис. 9.2. Процесс выполнения вдоха: а — объем грудной клетки в покое; б — увеличение объема при мышечном сокращении; в — увеличение обеспечивает снижение давления в легких и устремление воздуха в них положение. В результате расслабления внешних межреберных мышц ребра и грудина опускаются вниз, занимая обычные для состояния покоя положения. Как только это происходит, легочная ткань также принимает положение, характерное для состояния покоя. Как следует из рис. 9.3, 6, это приводит к повышению давления в грудной клетке, вследствие чего из легких выходит воздух. Выдох завершен. При дыхании с усилием выдох становится более активным процессом. Внутренние межреберные мышцы более активно тянут ребра вниз. Им могут помогать широчайшая мышца спины и поясничная квадратная мышца. Сокращение мышц живота повышает внутрибрюшное давление, вызывая движение внутренних органов вверх к диафрагме и ускоряя ее возврат в исходное дугообразное положение. Эти мышцы, кроме того, тянут грудную клетку вниз и вовнутрь. ДИФФУЗИОННАЯ СПОСОБНОСТЬ ЛЕГКИХ Газообмен в легких, который называется диффузией, выполняет главные функции: 1) восполняет запасы кислорода в крови, истощающиеся на тканевом уровне, при его использовании для образования энергии путем окисления; 176
Рис. 9.3. Процесс выполнения выдоха: а — объем грудной клетки возвращается к объему, характерному для состояния покоя; б — воздух устремляется из легких наружу Изменение внутрибрюшного и внутригрудно-го давления не только способствует выполнению дыхания с усилием, но и обеспечивает возврат венозной крови в сердце. С увеличением давления оно передается в большие вены, по которым кровь транспортируется к сердцу через брюшной и грудной участки. При снижении давления размер вен возвращается к исходному и они наполняются кровью. Изменение давления в брюшном и грудном участках вынуждает кровь идти в вены, обеспечивая ее возврат с помощью " выдаивающего" действия. В этом заключается сущность венозного возврата. Сокращения мышц при выполнении физической нагрузки также обеспечивают подобное " выдаивающее " действие, способствуя венозному возврату 2) выводит диоксид углерода из венозной крови. Для осуществления диффузии необходимы воздух, транспортирующий кислород в легкие, и кровь, утилизирующая этот кислород и выделяющая диоксид углерода. Воздух поступает в легкие во время легочной вентиляции, при этом осуществляется газообмен между ним и кровью. В ОБЗОРЕ... 1. Легочная вентиляция (дыхание) представляет собой процесс, обеспечивающий поступление воздуха в легкие и выведение его из них. Она состоит из двух фаз — вдоха и выдоха. 2. Вдох — активный процесс, во время которого увеличиваются размеры диафрагмы и внешних межреберных мышц, а следовательно, и объем грудной клетки. Это приводит к снижению давления в легких и поступлению в них воздуха. 3. Выдох, как правило, — пассивный процесс. Дыхательные мышцы расслабляются, а эластичная ткань легких принимает исходное положение, обеспечивая возвращение объема грудной клетки в обычное исходное положение. Это приводит к увеличению давления в легких и выделению из них воздуха. 4. Вдох и выдох, выполняемые с усилием, — активные процессы, зависящие от мышечных сокращений. Кровь из большинства участков тела возвращается по полой вене в легочную (правую) часть сердца. Из правого желудочка она выкачивается через легочную артерию в легкие, прокладывая 177 себе путь к легочным капиллярам. Эти капилляры образуют плотную сеть вокруг альвеолярных мешочков. Легочные капилляры настолько малы, что эритроциты могут передвигаться по ним лишь в один ряд, подвергаясь воздействию окружающей легочной ткани. Именно здесь происходит процесс диффузии. ЛЕГОЧНАЯ МЕМБРАНА Газообмен между воздухом в альвеолах и кровью в легочных капиллярах осуществляется через легочную мембрану (или как ее еще называют, — альвеолярно-капиллярную мембрану). Эта мембрана (рис. 9.4) состоит из альвеолярной стенки; капиллярной стенки и их базальных мембран. Респираторная мембрана очень тонкая — 0.5 — 4.0 мкм. Вследствие этого приблизительно 300 млн альвеол находятся в непосредственной близости от циркулирующей по капиллярам крови. Тем не менее эта мембрана является значительным препятствием для осуществления газообмена. Рассмотрим, как он происходит. ПАРЦИАЛЬНОЕ ДАВЛЕНИЕ ГАЗОВ Воздух, которым мы дышим, представляет собой смесь газов. Каждый из них оказывает давление, пропорциональное его концентрации в этой смеси. Давление каждого газа в смеси называется парциальным давлением. Согласно закону Дальтона, общее давление смеси газов равно сумме парциальных давлений каждого газа в этой смеси. Рассмотрим состав воздуха, которым мы дышим. Он состоит на 79, 04 % из азота (М^), на |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Последнее изменение этой страницы: 2019-06-19; Просмотров: 444; Нарушение авторского права страницы