|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ГРАНИЦЫ ПРИМЕНЕНИЯ ГИПОТЕЗЫ АНАЛИЗА ПРИЗНАКОВ
Почему же неправильно рассматривать перцептивные процессы как безобразный анализ признаков? По трем причинам. Во-первых, анализируемые признаки не являются какими-то особыми, отличными от других. Во-вторых, богатство феномена восприятия не объясняется теми детекторами признаков, которые открыты. И в-третьих, изменения, которые имеют место в системах входа в ходе развития организма, позволяют различать эффекты, возникающие в детекторах признаков, изученные с помощью микроэлектродов, и эффекты, продуцируемые в системе, анализирующей эти признаки, что было изучено в поведенческих опытах с различением сигналов. Рассмотрим прежде всего проблему различимости признаков. И. Рок (1970) со всей проницательностью сформулировал эту проблему следующим образом: «... Те, кто утверждал бы, что ответ нейронных детекторов может объяснить, почему вещи выглядят такими, какими они являются на самом деле, возможно, не знают тех фактов, касающихся восприятия, на которые гештальтисгы обращали особое внимание. Например, известно, что воспринимаемая нами величина, не просто зависит от величины образа на сетчатке, воспринимаемая форма не является непосредственной функцией формы соответствующего изображения, а воспринимаемая скорость — непосредственной функцией скорости перемещения изображения на сетчатке и т. д. Напротив, несмотря на значительные изменения находящегося в отдалении от нас стимула, возникающие при изменении позиции наблюдателя, предметный мир сохраняет удивительную константность. Эти факты нельзя отрицать, и их нельзя оставить без внимания на том основании, что вещи в этих изменяющихся условиях не выглядят константными, а мы знаем, что они константны. Против этой формулировки имеются достаточно веские возражения. Рассмотрим детально некоторые факты, касающиеся восприятия движения. Начнем с того, что существует константность положения, а именно-известно, что, когда мы перемещаемся или двигаем глазами, перемещается и образ всей картины на сетчатке, но сама картина не кажется движущейся. Случается также обратное, а именно неподвижный образ на сетчатке вызывает восприятие движения при движении наших глаз или го- ловы, как это наблюдается при слежении глазами за движущимся объектом или при рассматривании последовательного образа во время движения тлаз. Известно, далее, стробоскопическое движение, когда сменяющиеся неподвижные изображения вызывают ощущение движения. Известен также •феномен индуцированного движения предмета, изображение которого на сетчатке неподвижно. Этот феномен возникает вследствие перемещения окружающих объектов. Так, например, Луна кажется движущейся на ■ фоне перемещающихся облаков. Можно принять гипотезу, что действительной сенсорной основой восприятия движения является стимуляция детекторов движения, но тогда необходимо предположить, что значимость такой стимуляции определяется другой информацией. Например, можно сказать, что движение вообще не воспринимается, если данным о смещении изображения на сетчатке противостоит информация о том, что оно возникает в результате собственного произвольного движения организма, как это предположил фон Хольст, -а до него Гельмгольц. Или можно считать, что в случаях индуцированного движения детекторы предоставляют решающую информацию о том, что что-то движется, но по неизвестным еще причинам движение приписывается неподвижному объекту. Откровенно говоря, я не считаю это достаточно убедительным. Это не объясняет, почему я вижу свой последовательный образ движущимся, когда я двигаю глазами даже в совсем темной комнате, где не видно неподвижных предметов, изображения которых могли бы перемещаться на сетчатке. В случае же стробоскопического движения, если наши глаза удерживаются в стационарном положении, то источником стимуляции детекторов движения мог бы быть только возникающий в этих условиях нистагм глаза. Но это не могло бы объяснить движения, которое возникает между двумя раздельно освещаемыми объектами. Эта гипотеза не может объяснить и индуцированного движения, потому что предмет, который вызывает ощущение движения, может двигаться так медленно, что его движение будет ниже порога, в то время как только оно и воспринимается. Далее, очевидно, что в данном случае основой восприятия движения является изменение локализации предмета, который кажется движущимся относительно местонахождения предмета, который вызывает это движение, а не стимуляция детекторов перемещения изображения. Рассмотрим теперь следующую проблему — проблему восприятия ориентации предмета. Является ли основой воспринимаемой ориентации линии ориентация ее изображения на сетчатке? Очевидно, нет, поскольку при повороте головы вертикальные и горизонтальные линии на стене продолжают оставаться для нас вертикальными и горизонтальными. Здесь мы «нова сталкиваемся с константностью, в данном случае с константностью восприятия направления, несмотря на изменения в ориентации изображений на сетчатке. Это происходит в значительной степени даже тогда, когда мы рассматриваем в темноте одну светящуюся линию, наклонив или откинув голову в другую сторону. Следовательно, обоснованно заключение, что, прежде чем сделать вывод о значении для восприятия ориентации данного сетчаточного образа, необходимо принять во внимание информацию о собственной позиции наблюдателя. Далее, как бы мы поняли открытие, сделанное Хьюбелом и Визелом, о существовании в зрительной коре детекторов ориентации? Очевидно, что они не могут быть знаками ориентации объектов в окружающей среде. Можно согласиться, что они являются знаками, указывающими на ориентацию объектов относительно организма, другими словами, детекторами эгоцентрической ориентации, и что они создают условия, необходимые для того, чтобы организм мог определить ориентацию объектов в окружающей среде, когда он дополнительно получает информацию и об ориентации самого ' наблюдателя. Например, можно сказать, что, когда изображение линии на сетчатке будет иметь вертикальное положение, оно всегда будет казаться эгоцентрически вертикальным, то есть параллельным продольной оси головы, и линия будет восприниматься вертикальной в поле зрения наблюдателя, если он занимает вертикальное положение, и горизонтальной, если наблюдатель повернут на 90°. Хотя такое объяснение кажется мне правдоподобным, оно вызывает серьезные трудности из-за существования некоторых дополнительных фактов. Если наблюдатель сохраняет вертикальное положение, то вертикальная линия, воспринимаемая им на фоне изображения комнаты, при наклоне последнего будет выглядеть наклоненной в направлении, противоположном наклону изображения комнаты. Таким образом, в данном случае вертикальное изображение на сетчатке не будет более соответствовать вертикально расположенному предмету, несмотря на то, что наблюдатель находится в вертикальном положении. Более того, линия не будет уже казаться эгоцентрически вертикальной линией. К этим фактам можно было бы добавить данные о том, что наблюдатели будут адаптироваться к повернутым с помощью призматических стекол изображениям, так что вертикальное изображение на сетчатке не будет больше ни знаком вертикального объекта, когда наблюдатель находится в вертикальном положении, ни фактически знаком того, что объект ориентирован в вертикальном направлении относительно наблюдателя. Имеют ли какое-либо отношение детекторы ориентации в коре к восприятию формы? Можно предположить, что восприятие формы сводится к восприятию ориентации линий, из которых строится форма. Таким образом, предмет воспринимается как квадрат тогда, когда одновременно стимулируются два детектора вертикальных и два детектора горизонтальных линий. Хорошо известно, что фигуры выглядят весьма по-разному, когда меняется их ориентация. На первый взгляд сам этот факт может казаться доказательством того, что детекторам ориентации принадлежит существенная роль в восприятии формы, потому что в этом процессе принимают; участие совершенно различные волокна. Можно сказать, что это является основой для понимания явления изменения внешнего вида предмета. Однако в действительности, вообще говоря, объяснением изменения внешнего вида предмета служит не изменение ориентации изображения на сетчатке. В проведенных мною и моими сотрудниками экспериментах неоднократно-было показано, что, если положение фигуры в отношении ее окружения не меняется, но она рассматривается наблюдателем в наклонной позе, фигура не выглядит измененной, или, выражаясь более точно, распознавание не ухудшается. Могло ли бы это быть, если за это явление были бы ответственны совершенно различные детекторы? Напротив, если фигура наклонена так же, как и наблюдатель, и ориентация изображения на сетчатке, таким образом, совсем не изменилась, фигура тем не менее выглядит совершенно иной и иоэтому часто не узнается. Можно провести простой опыт: создайте последовательный образ квадрата, держа голову в вертикальном положении, а затем, закрыв глаза и повернув голову на 45°, посмотрите на него, он будет подобен ромбу. Могло ли бы это быть, если за это явленяе были бы ответственны одни и те же детекторы? Однако эти факты приобретают смысл, если считать, что решающим-для ориентации является восприятие определенной части фигуры как верх, а другой — как низ и т. д. Если мы будем правильно оценивать одну и ту же часть фигуры в качестве верха, как мы это обычно делаем, приняв наклонное положение, то мы увидим, что форма предмета не меняется. Однако, если мы за верх фигуры принимаем другую ее часть, как это бывает при наклоне фигуры относительно окружения, то мы увидим, что ре форма очень изменилась. Этот процесс, по-видимому, не имеет ничего-общего со специфическими детекторами ориентации. Далее, роль этих детекторов, возможно, состоит в том, что они служат для различения ориентации одного образа по отношению к другому. В прошлом мы ошибочно считали, что такое различение осуществляется благодаря различной пространственной ориентации проекционных кортикальных «образов». Иными словами, нейронные детекторы, вероятно, яв-
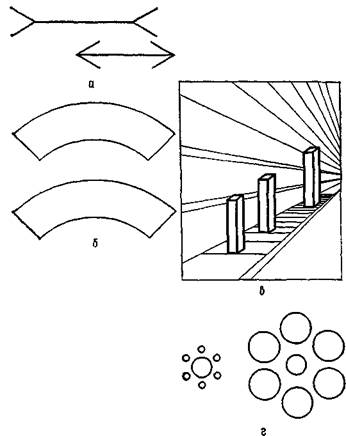
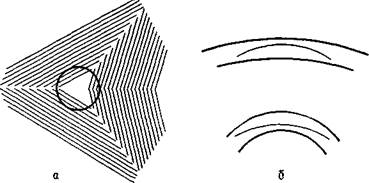
Рис. VII-10. Некоторое нарушение константности величины (иллюзии) при тождественности сетчаточных образов. а — классическая иллюзия Мюллера — Лайера, когда две одинаковые линии кажутся различными из-за стрелок на их концах, которые направлены в противоположные стороны; б — нижняя фигура кажется больше верхней, хотя обе имеют один и тот же размер; в — три колонки имеют одинаковую высоту; г — два центральных круга равны. (Kretch and Crutchfield, 1962). ляются механизмами, посредством которых воспринимается определенная информация о стимуле, в данном случае информация о том, что сохраняется или изменяется пространственная ориентация. Конечно, такая информация необходима для восприятия в качестве исходной, но ответ волокон детекторов никоим образом нельзя рассматривать как нейронные корреляты единиц восприятия» (1970, р. 2—5). В остальной части статьи Рока анализируется еще ряд интересных экспериментов, которые трудно объяснить с позиции детекторов признаков (см. также рис. VII-10, VII-11). Второй недостаток точки зрения, рассматривающей восприятие как безобразный процесс выделения признаков, состоит в том, что по сравнению с богатством явлений сознания процесс распознавания структуры, осуществляемый методом классификации, довольно беден деталями, которыми он оперирует. Механизм распознавания зрительной структуры, который осуществляется путем избирательного выделения признаков, может приводить к ошибкам. Каждый из нас ждал кого-нибудь, кто опаз-
Рис. VII-11. Два примера, иллюстрирующие влияние окружающего фона на восприятие формы, а — фигура, лежащая на штриховом фоне, имеет форму точного круга; б—дуги, находящиеся внутри двух параллельных кривых, идентичны. Эти иллюзии трудно объяснить с позиции механизма детекции признаков (Kretch and Crutchfield, 1962). дывал на свидание, назначенное в многолюдном месте, и ошибочно принимал посторонних за ожидаемое лицо, так как они имели сходные с ним черты. Мы узнаем А, Л и а как букву А; мы хорошо знаем о различиях почерков. Восприятие, по-видимому, обеспечивает в одно и то же время не только богатство нашего опыта и «сильное предпочтение сохранять постоянную организацию», но также и тенденцию к «максимальному упрощению», как показали опыты гештальтпсихологов. Имеются данные, свидетельствующие о том, что богатство восприятия связано со сложностью конфигураций окружающей среды, которые существуют независимо от организма и возбуждают его рецепторные поверхности (см. Gibson, 1966), последние приводят в действие некоторый механизм мозга с соответствующей емкостью, позволяющей вместить это богатство. Данные, представленные Роком и другими, показывают, что только механизм выделения признаков обусловливает тенденцию к упрощению, создает схему отно-
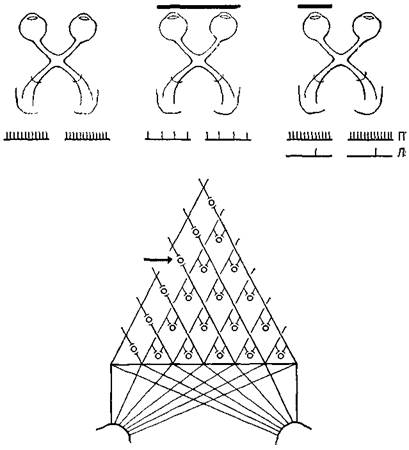
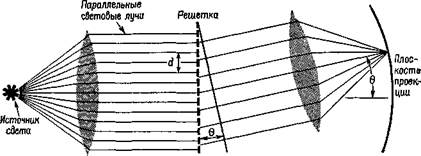
Рис. VII-12. Схема, иллюстрирующая результаты экспериментов со зрительной депривацией одного или двух глаз у новорожденных. Следует отметить, что односторонняя депривация ведет к значительно большему подавлению активности нейронов зрительной коры, чем двусторонняя. Это обстоятельство создает известные трудности (см. текст) при объяснении некоторых экспериментальных фактов с позиции простого механизма детекции признаков, поскольку нейроны имеют связи с обоими глазами (нижняя схема) и продолжают нормально отвечать на стимуляцию глаза, не подвергшегося сенсорной депривации. шений. Однако даже здесь начинает играть роль другой механизм — более сложный «познавательный» процесс. Третий недостаток объяснения фактов восприятия с позиции выделения признаков выступает в экспериментах, в которых новорожденные в течение различных периодов времени подвергались сенсорной депривации. Как уже говорилось в гл. VI и VIII, младенцы в возрасте нескольких недель обнаруживают способность к восприятию константности величины и формы. У животных способность к различению развивается также рано; при изучении мозга детенышей животных были обнаружены детекторы признаков, которые уже достигли своего полного развития. Однако при ограничении потока сенсорных воздействий эта простая корреляция между поведением и функциями мозга нарушается. Так, когда кошки и крысы содержатся в полной темноте, их способность различать яркость и контур остается неизмененной, но способность различать более сложные зрительные структуры значительно ухудшается. Хотя ориентация рецептивных полей кортикальных нейронов у таких животных еще различима, она имеет менее острую настройку, и утомление клеток развивается быстрее, чем у животных, содержащихся в нормальных условиях. Но когда сенсорная депривация ограничена депри-вацией одного глаза в течение определенного времени, как зрительная ориентация, так и способность к различению оказываются резко нарушенными, если животное пользуется только одним этим глазом. Это связано с тем, что стимуляция глаза, ■ подвергнутого депривации, может привести в активное состояние лишь небольшое число нейронов, которые вряд ли могут сколько-нибудь избирательно отвечать на определенным образом ориентированную линию или движение. Эта реакция возникает, даже несмотря на то, что в коре присутствует обычный для нее комплект клеток с их нормальной реактивностью, как это было обнаружено при стимуляции не подвергавшегося депривации глаза (см. рис. VII-12). Таким образом, когда имеет место нарушение в привычном способе возбуждения, происходит какое-то подавление реактивности и это подавление переходит в нарушение функции исключительно в результате неупотребления нервных структур (как у животного, подвергшегося полной сенсорной депривации). Если монокулярное подавление реактивности коры объясняется некоторым изменением в связях, то ответственным за этот эффект подавления должен быть какой-то процесс, отличный от простого выделения признаков. Л. Ганц в недавнем обзоре (1971) этих и других сходных данных доказал это. Основываясь на результатах экспериментов с полной депривацией животных, он различает два типа заданий. Ориентация по яркости и контуру требует только, чтобы организм отвечал на языке реакций, свойственных большинству его детекторов признаков. Различение более сложных структур, например двух треугольников, один из которых перевернут, не может осуществляться на этой основе. Мы нуждаемся в каком-то дополнительном, более гибком мозговом механизме, который Ганц называет «избирательным вниманием». Этот механизм активно подавляет доминирующую реакцию детекторов признаков, реак- цию, которая, в сущности, не имеет отношения к адекватному решению задачи на различение. Следовательно, мы снова сталкиваемся с необходимостью иметь дополнительный механизм, помимо выделения признаков. Поэтому мы снова приходим к мысли о той, что, возможно, системы входа организованы таким образом, что нейронные сигналы координируются с каким-то психологическим процессом формирования образов. Проблема состоит в том, чтобы определить нейро-логическую природу этой организации. На основе рассмотренных в начале этой главы данных становится очевидно, что эта организация не может быть просто мозаикой точек, которые генерируются в рецепторах и без изменения проводятся до коры, где они формируют образ, подобно фотографии. Столь же очевидно, что следует принять во внимание наличие нейронов, чувствительных к линиям, и других детекторов признаков, которые создают в мозгу нейронную основу для их анализа. РЕЗЮМЕ Почти полное удаление мозговой ткани независимо от ео л№-кализации в пределах систем входа оказывает лишь незначительное влияние на процесс распознавания. Следовательно, логика работы нейронов, свойственная любому конкретному восприятию, распространяется, по-видимому, на всю систему. Были получены также прямые данные, касающиеся распределения памяти в мозгу. Логику этой памяти создают как встроенные детекторы элементарных признаков (таких, как линии и углы), так и анализаторы для более сложных преобразований особых признаков (таких, как волновые формы), которые перестраиваются в ходе жизненного опыта организма. Глава VIII Голограммы Гипотеза Вернемся на время к нашей общей модели. Модель предполагает два основных процесса: пространственно организованные-состояния и операции, выполняемые на фоне этих состояний с помощью импульсной передачи между нейронами. В первой части книги было описано, каким образом эти основные свойства нейронных групп могут комбинироваться в логические операции, усиливающие аналитические и контрольные (серво) функции" нервной системы. Учитывая их значение, а также то, что сейчаа они игнорируются в нейропсихологической и нейрофизиологической литературе, я подчеркнул, что построение имеющих определенную структуру топологических, то есть пространственных, представительств в нервной системе является одной из форм, которые могут принимать состояния мозга. Я предположил, что-взаимодействие динамических структур возбуждения, падающих на рецепторные поверхности, после их передачи по параллельным путям кодируется благодаря горизонтальным связям в активность медленных потенциалов групп нейронов и образует временные микроструктуры, рисунки которых зависят скорее от функциональной организации нейронных соединений, чем от нейронов, как таковых. В гл. VII были подробно рассмотрены факты, свидетельствующие о существовании детекции признаков и механизма анализа, а также приведены аргументы в пользу необходимости особой нейронной организации, помимо той, которая представлена анализаторами признаков. В гл. VIII формулируется гипотеза о том, что такой организацией может быть механизм микроструктуры медленных потенциалов соединительных аппаратов. Эта гипотеза основывается на предположении, что нейронное отображение входных воздействий не является фотографическим и создается не только посредством имеющейся системы фильтров, выделяющих признаки, но и с помощью особого класса преобразований, которые обладают значительным формальным сходством с процессом отражения оптического образа, открытым математиками и инженерами. Этот оптический процесс, названный голографией, основан на использовании явления интерференции структур. Он обладает множеством удивительных свойств, из которых первостепенное значение имеет его способность к распределению и сохранению большого количества информации. Именно эти свойства дают возможность разрешить противоречие между потребностью в функциональной лабильности, быстром темпе изменений и уже рассмотренными анатомическими особенностями в организации нервных систем приемл информации. Прежде чем точно сформулировать проблему, полезно посвятить несколько параграфов разъяснению нашего общего подхода. Обработка оптической информации с помощью голографии описывается математически в понятиях механической волны. В физической оптике для описания распространения света в экспериментальных условиях могут быть использованы как квантовые, так и волновые уравнения. Физика не интересует то, является ли свет квантом или волной, распространяется ли он в виде частиц или электромагнитных волн, или в форме того и другого. Он заботится о том, чтобы как можно полнее количественно описать результаты своих наблюдений, и соответственно этому выбирает инструменты для своего описания. Некоторые наблюдения могут быть хорошо описаны как статистическая вероятность квантовых событий, другие легче описываются с помощью волновой теории. Особенно трудно привести в соответствие с квантовым подходом одно наблюдение. Когда свет очень низкой интенсивности проходит через решетку, то есть когда предполагается, что через каждую отдельную щель пройдет самое большее несколько квантов света, уравнение, описывающее световой поток по другую сторону решетки, для объяснения наблюдаемых эффектов должно учитывать некоторое взаимодействие, которое, по-видимому, имеет место между световыми потоками, проходящими через соседние щели (рис. VIII-1). Трудно, хотя и не невозможно, объяснить это взаимодействие с точки зрения квантовой теории: каким образом квант света может влиять на соседние, отделенные от него решеткой? Кажется, будто каждый квант оказывает силовое (возникает искушение сказать «магнетическое») действие на соседний. Этот феномен взаимодействия был широко изучен и получил название суперпозиции. Математические уравнения, удовлетворительно объясняющие явление суперпозиции, представляют собой систему линейных уравнений, названных интегралами свертки, которые обычно используются для описания взаимодействия волновых процессов. Описание одной волновой формы свертывается вместе с описанием другой. При объяснении суперпозиции не возникает серьезных теоретических проблем,
Рис. VIII-1. Дифракционная решетка (Murray and Cobb, 1970;. если считать, что ровный фронт волн света, встречаясь с решеткой, разбивается ею на небольшие, взаимодействующие между собой «вихри». Можно считать, что такие вихри и вызывают эффект интерференции и объясняют суперпозицию — «взаимодействие по соседству». Голографическая гипотеза функционирования мозга в процессе восприятия принимает форму суперпозиции. Фактически выбор способа объяснения того, как изменяются электрические потенциалы мозга — в статистических терминах или в волновых, — зависит от наблюдений, которые должны быть описаны. Следует выбрать тот способ, который может дать наиболее полное количественное описание наблюдений и в то же время быть приемлемым для концепции. В гл. III описано возникновение латерального, или периферического, торможения в рецепторах и на разных уровнях центральной нервной системы. Латеральное торможение обусловливает эффект взаимодействия между процессами, протекающими в соседних нейронах. Если это пространственное взаимодействие адекватно описывается интегралами свертки, которые характеризуют физическое явление суперпозиции, то мы можем рассматривать взаимодействие в нервной системе на основе интерференционных эффектов. Правомочность такого выбора подтверждается теми фактами, что по крайней мере в сетчатке взаимодействие между соседними горизонтальными клетками обусловлено исключительно тормозными взаимодействиями посредством медленных потенциалов гиперполяризации, а не деполяризации, которая приводит к возникновению нервных импульсов. Как и в гл. III, рецепторные явления служат миниатюрными моделями на этот раз «нейронного голографического» процесса. Мы уже отмечали, что возбуждение одной единицы зрительного нерва оказывает влияние на частоту разрядов соседних
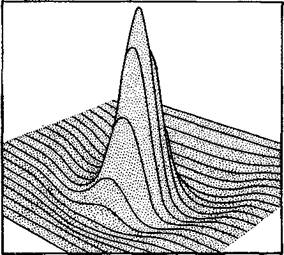
Рис VIH-2 Карта рецептивного зрительного поля одиночной ганглиозной клетки сетчатки, построенная на основе записей активности ее аксона при точечном световом раздражении различных участков зрительного поля Карта представляет собой семейство контурных линий, так как ганглиозная клетка интегрирует ответы соседних нейронов, с которыми она связана, см. рис Ш-4 Высота контурной линии в любой ее точке дает представление о числе нейронов, которые одновременно активируются при предъявлении точечного раздражения в данном пункте рецептивного поля Максимум нейронных разрядов возникает, когда расположение светового пятна соответствует позиции центрального пика. В математическом смысле каждая контурная линия представляет собой «интеграл свертки» первой производной от формы светового раздражителя Взаимодействие многих таких интегрэлов свертки может вызвать в зрительной системе и в любом друюм месте мозга интерференцию динамических структур, подобную топографической Сохранение таких структур могло бы быть основой для системы распределенной памяти (Rodieck, cm Pribram, 1969). «единиц. Мы отметили также, что рецептивное поле отдельной единицы образуется в результате такого пространственного взаимодействия между соседними элементами. В зрительном нерве эти рецептивные поля обычно состоят из более или менее округлого центрального пятна, которое реагирует либо возрастанием (on — -Центральный нейрон), либо снижением (off — центральный ней- рон) частоты своих спонтанных разрядов, и из окружающей зоны, которая характеризуется активностью, противоположной по знаку активности центра. Глава VII была посвящена проблеме: .какой тип кода, какой тип организации создается в результате действия многих таких единиц? Настоящая глава посвящена другому кругу проблем, а именно: что кодируется в одном рецептивном поле, то есть какие черты оптического изображения определяют форму рецептивных полей? Ответ на этот вопрос следует из проделанного Родие-ком (1965) детального математического анализа количественных отношений, существующих между специфическим образом движущимися зрительными стимулами (вспышками) и соответствующими им формами рецептивного поля. Кривые полученных ответов (каждая из которых представляет собой вертикальный срез через трехмерное рецептивное поле, рис. VII1-2) имеют особую 'форму. При одновременном предъявлении двух или более стимулов кривая ответов достаточно хорошо соответствует кривой, которая возникает в результате суперпозиции кривых ответов на раздельные предъявления этих стимулов. Таким образом, Родиек делает зывод, что, исходя «из структур ответа на небольшое световое пятно, возникающих при засвечивании разных участков рецептивного поля клетки, можно получить структуру ответа этой клетки на фигуру любой формы, движущуюся в любом направлении, с любой скоростью, через любую часть рецептивного поля». Это открытие позволило воспроизвести рецептивное поле клетки, используя только движущиеся пятна света, а также сделало возможным с помощью компьютера произвести запись, показанную на рис. II1-7. Таким образом, форма рецептивного поля оказывается сверткой первой производной от формы зрительного стимула. Как уже отмечалось, интегралы свертки, устанавливающие соотношения между событиями, происходящими в соседних областях (пространственная суперпозиция), описывают основной го-лографический процесс в системах, перерабатывающих оптическую информацию (Gabor, 1949, 1951). Эффекты интерференции ■ описываются также уравнениями преобразований Фурье, Френеля и Лапласа. Определение же того, какое преобразование больше подходит для описания действия системы в данных условиях, является эмпирической проблемой. В зрительной системе — и имеются достаточные основания благодаря работам Г. фон Бекеши (1960, 1967, рис. VIII-3), чтобы предполагать, что также и в слуховой, соматической и вкусовой системах, — эти виды преобразований с большой точностью описывают взаимодействие между структурами энергетических воздействий, возбуждающими рецепторные поверхности, ж спонтанными изменениями потенциалов в рецепторных единицах. В системах, обрабатывающих оптическую информацию, такие преобразования обычно имеют отношение к анализу формы волны (то есть эффектов интерференции), поэтому нет особых причин полностью игнорировать волновой подход и при объяснении эффекта суперпозиции, возникающего в нейронных системах. Преимущество такого подхода состоит в том, что физический процесс, основывающийся на эффектах интерференции, обладает-
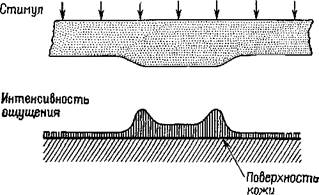
Рис. VIII-3. Схема эксперимента, показывающего существование полос Маха для кожной чувствительности В эксперименте использовалось вибрационное точечное раздражение, обозначенное на схеме как стимул. Ощущение, вызываемое вибрационным раздражением, приложенным к предплечью, обозначено на схеме как интенсивность ощущения. Сходный, но несколько ослабленный эффект можно получить в результате прямого давления (с различной интенсивностью) невибрирующего стимула (Bekesy, 1967). многими свойствами нервного процесса восприятия и, таким образом, облегчает понимание мозгового механизма формирования образов. Рассмотрим теперь физическую голограмму. ФИЗИЧЕСКАЯ ГОЛОГРАММА Большинству из нас известно, каким образом возникает изображение в обычных оптических системах. Фотоаппарат регистрирует на фотопленке, помещенной в плоскости изображения, копию пространственного распределения интенсивности света, отраженного от предметов, попавших в поле объектива. Каждая точка на фотопленке сохраняет информацию, полученную от соответствующей точки зрительного поля, и, таким образом, зарегистрированная картина выглядит подобно картине зрительного поля. Недавно-были проведены исследования свойств записей, сделанных на
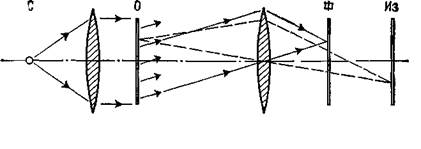
Монохромапш- Обьект Фильтр Изобра- ческий точечный б фокальной жение источник света плоскости Рис VIII-4. Схематическое представление метода, используемого для создания оптического топографического фильтра (van Heerden, 1968). фотопленке, которая находится в оптической системе где-то перед плоскостью изображения (то есть в фокальной плоскости). При соответствующей экспозиции такой фотопленки с помощью источника когерентного света она становится оптическим фильтром, в котором информация о каждой точке зрительного поля распределяется и хранится по всему фильтру (рис. VIII-4). Такие фильтры обладают многими удивительными свойствами. Как мы знаем, когда фотопленка внутри камеры находится не точно в плоскости оптического изображения, образ становится расплывчатым, его границы менее четки, контраст менее выражен. В оптическом фильтре информация распределяется таким образом, что зафиксированное на пленке изображение вообще не является подобием зрительного изображения. Оптический фильтр представляет собой запись волновых узоров, излучаемых объектом или отражаемых от него. «Такую запись можно рассматривать как «замороженный» волновой узор, который остается «замороженным» до тех пор, пока не приходит время восстановить процесс, после чего волны «считываются» с регистрирующего посредника» (Leith and Upatniecks, 1965). Так, когда оптический фильтр освещается источником когерентного света, он восстанавливает волновые фронты света, которые имели место при экспозиции объекта. В результате при рассматривании ■ фильтра становится видно действительное изображение зрительного поля. Этот зрительный образ, по-видимому, точно совпадает ■ с картиной, имевшей место при экспозиции, он является ее пол-вым трехмерным изображением (рис. VIII-5). По существу, вся информация, описывающая зрительное поле, из которой может быть восстановлено изображение, содержится в этом фильтре. «Когда наблюдатель при рассмотрении фильтра меняет свое положение, перспектива изображения тоже меняется, как если бы наблюдатель рассматривал реальную сцену. Обнаружены эффекты параллакса между близко и далеко расположенными объектами: если один предмет находится на переднем плане и заслоняет какой-то другой предмет, то наблго- датель может, двигая головой, заглянуть за заслоняющий предмет и: таким образом увидеть то, что за ним скрывается... Короче говоря, восстановленная голограмма обладает всеми визуальными свойствами реальной сцены, и мы не знаем ни одного зрительного теста, который можно было бы использовать, чтобы отличить их друг от друга (Leith and Upatnieeks,. 1965, p. 30). Еще до того, как было продемонстрировано практически использование оптических фильтров в восстановлении изображений, Д. Габор (1949, 1951) математически описал другой способ-
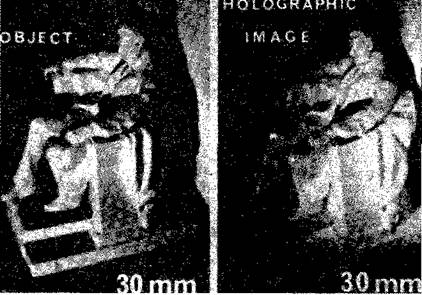
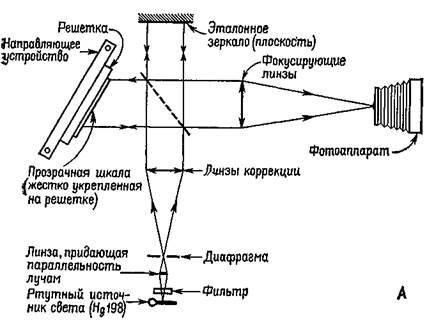
Рис. VIII-5. Обычная фотография объекта и фотография реального изображения того же самого объекта, сделанная голографическим методом (справа), обе с помощью лазерного источника света. Обратите внимание,, что при современной технике голографическое изображение достигает высокого уровня совершенства (Stroke, 1969). получения изображения с помощью фотографии. Габор стремился увеличить разрешающую способность электронной микрофотографии. Он предположил возможность интерференции когерентной фоновой волны с волнами, которые преломляются тканью. (Этой цели может служить также отражение волн от темного объекта.) Возникающий в результате интерференции^ узор сохранял бы и амплитудную и фазовую (взаимодействие^ по соседству) информацию, которая затем на втором этапе могла. JP и с. VIII-6. А — схема, иллюстрирующая метод получения голограмм по Габору; Б — фотография аппаратуры (Stroke, 19G9).
бы быть использована для восстановления при освещении источником когерентного света изображения естественной ткани. Габор-назвал свою технику «голографией», а сфотографированную картину— «голограммой», потому что она содержала всю информацию, необходимую для восстановления целостного образа. Голограммы Габора можно построить двумя способами. Волна разделяется с помощью прибора, расщепляющего лучи (то есть полупрозрачного посеребренного зеркала) таким образом, что-одна часть служит в качестве опорной волпы, а другая отражается от объекта, который должен быть сфотографирован (рис. VIII-6r VIII-7). Затем можно использовать один опорный луч для восстановления изображения. Либо каждая часть расщепленного-луча может быть отражена от различных объектов. Когда при восстановлении изображения один из объектов используется в качестве опорного, другой появляется как образ его «призрака» (фантомное изображение). В этом случае голограмму можно использовать как механизм ассоциативного хранения информации. Популярное:
|
Последнее изменение этой страницы: 2016-03-17; Просмотров: 924; Нарушение авторского права страницы