|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ. ПАРАМЕТРЫ ВОЗБУДИМОСТИ. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ
Актуальность темы (мотивация). В процессе эволюции происходила постоянная дифференцировка тканей, участвующих в приспособительной деятельности организма. Ткани, способные с максимальной скоростью, целенаправленно отвечать на раздражение активной специфической реакций, получили название возбудимые (мышечная, нервная, железистая). Возбудимость и ее изменения определяют самые различные стороны деятельности организма, в результате обеспечивает наиболее оптимальное приспособление организма к изменяющимся условиям окружающей среды. Иногда необходимо воздействовать на возбудимость тканей с лечебной целью. Изучение электрических явлений, сопровождающих процессы покоя, возбуждения и торможения в живых тканях, имеет большое значение для понимания природы этих процессов и для выявления характера нарушений деятельности возбудимых клеток при различных видах патологии.
Дидактические (учебные) цели: - дать представление о физиологических свойствах возбудимых тканей, методах раздражения возбудимых тканей, законах раздражения, параметрах возбудимости; - ознакомить с понятиями потенциал покоя, потенциал действия, дать представление о современных теориях происхождения мембранного потенциала и потенциала действия; - дать представление о местном и распространяющемся возбуждении. · Студент должен знать: - виды возбудимых тканей, их физиологические свойства, методы раздражения возбудимых тканей, законы раздражения, параметры возбудимости; - современные представления о происхождении биоэлектрических явлений в живых тканях, уравнения Нернста и Гольдмана; - понятия потенциал покоя и потенциал действия, фазные изменения возбудимости при генерации потенциала действия; - особенности местного и распространяющегося возбуждения.
· Студент должен уметь: - обездвижить, зафиксировать лягушку, приготовить нервно-мышечный препарат; - определить реобазу и конституционную хронаксию при прямом и непрямом раздражении на нервно-мышечном препарате лягушки; - определить реобазу и субординационную хронаксию локтевого нерва и мышц кисти у человека; - воспроизвести первый и второй опыты Гальвани; - воспроизвести опыт вторичного тетануса Маттеучи; - зарегистрировать потенциал повреждения скелетной мышцы у лягушки; - объяснить принципы регистрации потенциалов действия мышцы у человека методом электромиографии; Вопросы для самоподготовки I. Вопросы по базисным знаниям 1. Строение, свойства и основные функции биологических мембран. 2. Механизмы трансмембранного транспорта.
II. Вопросы по теме занятия 1. Виды раздражителей, особенности электрических раздражителей. 2. Физиологические свойства возбудимых тканей. 3. Законы раздражения возбудимых тканей. Параметры возбудимости. 4. Законы действия постоянного тока. 5. История открытия биоэлектрических явлений. 6. Мембранный потенциал. Происхождение и механизмы поддержания величины мембранного потенциала. 7. Потенциал действия. Критический уровень деполяризации. 8. Фазные изменения возбудимости при генерации потенциала действия. 9. Отличительные особенности местного и распространяющегося возбуждения.
III. Профильные вопросы
· Специальность: «Лечебное дело» 1. Применение постоянного тока в клинической практике. · Специальность: «Педиатрия» 2. Особенности потенциала покоя и потенциала действия в раннем онтогенезе. · Специальность: «Медико-профилактическое дело» 3. Методы оценки функционального состояния мышечной ткани при профосмотрах. · Специальность: «Медико-биологическое дело» 4. Методы изучения ионных каналов мембран. · Специальность: «Стоматология» 5. Электромиографические исследования в стоматологии. · Специальность: «Восточная медицина» 6. Оценка состояния возбудимых тканей методами восточной медицины. · Специальность: «Фармация» 7. Специфические блокаторы ионных каналов мембран. IV. Вопросы для самоконтроля 1. Структурные элементы плазматической мембраны. 2. Методы регистрации мембранных потенциалов. 3. Изменение проницаемости мембраны для ионов натрия при возбуждении. 4. Фазы потенциала действия. 5. Реполяризация мембраны. 6. Свойства локального ответа. 7. Распространяющееся возбуждение.
Краткое содержание темы (по вопросам для самоподготовки)
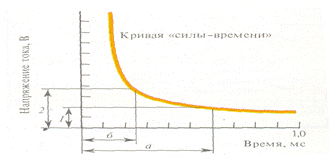
Виды раздражителей, особенности электрических раздражителей. По биологическому значению все раздражители разделяют на адекватные и неадекватные. Адекватным считается такой раздражитель, к восприятию которого данная биосистема специально приспособилась в процессе эволюции. Так, для органа зрения адекватно электромагнитное воздействие в определенном диапазоне длин волн; для слуха - упругие механические колебания среды и т.д. К категории неадекватных относят раздражителей, не являющихся в естественных условиях средством возбуждения данной биосистемы, но тем не менее, способных при достаточной силе вызвать возбуждение. В зависимости от силы действия на возбудимые ткани, раздражители подразделяются на пороговые, подпороговые, сверхпороговые. Минимальная сила раздражителя, необходимая для возникновения минимального по величине возбуждения, называется пороговой силой, или порогом возбуждения. По величине порога возбуждения оценивают возбудимость тканей, она у них различная. Раздражители, сила которых ниже порога возбуждения, рассматриваются как подпороговые. Сила раздражителя, вызывающая наибольший (максимальный) ответ ткани, называется максимальной силой раздражения, или сверхпороговой силой раздражения. Раздражители могут иметь физическую (электромагнитные волны, электрический ток, механические воздействия и т. д.), химическую (газы, различные химические соединения), физико-химическую (изменения ионного состава, рН, осмотического давления и т. д.), биологическую (вирусы, различные микроорганизмы и т.д.) природу. Электрические раздражители чаще всего используютсяв физиологических экспериментах, потому что они не оставляют в тканях необратимых явлений, легко дозируются по силе и длительности, по своей природе близки к электрическим процессам, которые возникают в возбудимых тканях. Физиологические свойства возбудимых тканей. В основе приспособительных реакций организма лежит универсальное свойство всех живых тканей – раздражимость, т.е. их способность отвечать на действие раздражающих факторов изменением структурных и функциональных свойств. Раздражимость является эволюционно древней формой реагирования недифференцированных тканей. В процессе эволюции произошла постепенная дифференцировка тканей, участвующих в приспособительной деятельности организма. Раздражимость в этих тканях достигла наивысшего выражения и получила название возбудимость. В физиологии к возбудимым тканям принято относить преимущественно мышечную, железистую и нервную ткани. Возбудимость – это способность тканей специализированно, целенаправленно и с максимальной скоростью отвечать на раздражение. Физиологическим проявлением процесса возбуждения у них является специализированная форма ответной реакции. Так, у мышечной ткани ответной реакцией на раздражение является сокращение, у нервной ткани – проведение электрического импульса, у секреторной ткани – синтез и выделение биологически активного вещества. Специфичный лишь для данного вида ткани ответ на раздражение получил название возбуждения. Возбуждение – сложный биологический процесс, характеризующийся изменением процессов обмена, теплообразования, временной деполяризацией мембраны клеток и другими физиологическими и биофизическими проявлениями. Невозбудимось тканей момент возбуждения получила название рефрактерность, различают абсолютную и относительную рефрактерность. Абсолютная рефрактерность – это невозбудимость ткани в момент возбуждения на любую силу раздражителя. Относительная рефрактерность – это возбудимость ткани в момент возбуждения на сверхпороговую силу раздражения. Для нервной и мышечной ткани характерно, что возбуждение, возникшее на одном участке мышечного или нервного волокна, может очень быстро передаваться на соседние участки того же волокна, т. е. они обладают проводимостью возбуждения. Н.Е. Введенский обратил внимание на разную способность возбудимых образований воспроизводить высокие ритмы возбуждений. Максимальное число потенциалов действия (''максимальный ритм''), которое способно возбудимое образование генерировать в 1 секунду, в соответствии с ритмом раздражения, он предложил в качестве показателя ''лабильности'' тканей. Предельный ритм импульсации в нервных и мышечных волокнах определяется скоростью изменения процессов ионной проводимости, лежащих в основе абсолютной и относительной рефрактерности. Все возбудимые ткани способны переходить с процесса возбуждения на процесс торможения. Законы раздражения возбудимых тканей . Параметры возбудимости. Закон силы утверждает, что с увеличением силы раздражителя возрастает величина ответной реакции ткани. Однако, величина ответной реакции ткани возрастает только до известного, определенного для каждого живого образования предела. Дальнейшее увеличение силы раздражителя уже не ведет к росту ответной реакции. В соответствии с этим законом функционирует скелетная мышца. Амплитуда ее сокращения постепенно увеличивается с увеличением силы раздражителя вплоть до достижения максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. На пороговые раздражители отвечают только волокна, имеющие самую высокую возбудимость, амплитуда мышечного сокращения при этом минимальна. Увеличение силы раздражителя приводит к постепенному вовлечению волокон, имеющих меньшую возбудимость, поэтому амплитуда сокращения мышцы усиливается. Когда в реакции участвуют все мышечные волокна данной мышцы, дальнейшее повышение силы раздражителя не приводит к увеличению амплитуды сокращения. Закон «Все или ничего» утверждает, что подпороговые раздражители не вызывают видимой ответной реакции тканей («ничего»), а на пороговые и сверхпороговые раздражители возникает одинаковая максимальная ответная реакция («Все»). По закону «Все или ничего» сокращаются сердечная мышца и одиночное мышечное волокно. Закон «Все или ничего» не абсолютен. Во-первых, на раздражители подпороговой силы не возникает видимой ответной реакции, но в тканях происходит изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа). Во-вторых, сердечная мышца, растянутая кровью, реагирует по закону «Все или ничего», но амплитуда ее сокращения будет больше, по сравнению с таковой при сокращении нерастянутой сердечной мышцы. Закон «силы-времени». Сила любого стимула в определенных пределах находится в обратной зависимости от длительности его действия на возбудимую ткань. Она имеет гиперболический характер (рисунок 1).
Рисунок 1. Кривая «силы – времени» 1 – реобаза; 2 – удвоенная реобаза а – полезное время действия тока; б – хронаксия
По этой кривой, прежде всего можно судить о том, что ток ниже некоторой минимальной силы, или напряжения, не вызывает возбуждения, как бы длительно он ни действовал. Чем больше ток, тем меньше времени ондолжен действовать на возбудимые ткани, чтобы вызвать возбуждение. Особенно четко эта зависимость проявляется при использовании в качестве раздражителя прямоугольных импульсов постоянного тока. Причиной такой зависимости является мембранная емкость. Очень «короткие» токи не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная сила постоянного тока, способная вызвать возбуждение была названа Л. Лапиком реобазой (порог возбуждения. Наименьшее время, в течение которого должен действовать раздражающий стимул величиной в одну реобазу, была названа полезным временем. Слово «полезное» было применено, с целью подчеркнуть, что дальнейшее увеличение длительности действия тока не имеет значения, т.е. бесполезно для возникновения потенциала действия. В качестве времени константы раздражения Л. Лапик ввел понятие «хронаксия» – это время, в течение которого должен действовать ток удвоенной реобазы, чтобы вызвать возбуждение. Закон градиента силы. Стимулирующее действие постоянного тока на возбудимые ткани зависит и от скорости нарастания тока во времени. При действии медленно нарастающего тока возбуждение не возникает, так как происходит приспособление возбудимой ткани к действию этого раздражителя, что получило название аккомодации. В основе аккомодации лежат инактивация натриевой и повышение калиевой проводимостей, развивающиеся во время медленно нарастающей деполяризации мембраны, в результате этого, происходит повышение критического уровня деполяризации. Способность к аккомодации у различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника и желудка. Законы действия постоянного тока. В 1859 году Э.Ф. Пфлюгер, изучая действие постоянного тока на ткани, установил ряд закономерностей, которые были сформулированы в виде законов. Полярный закон раздражения.Постоянный электрический ток обладает полярным действием на возбудимые ткани, т.е. он оказывает раздражающее действие на них в момент замыкания под катодом, а в момент размыкания - под анодом. При замыкании, когда возбуждение возникает под катодом, порог раздражения ниже, чем при размыкании, когда возбуждение возникает под анодом. Поэтому, при одной и той же силе тока раздражающее действие замыкания выражено сильнее, чем действие размыкания. Эти все факты именуются полярным законом раздражения Э.Ф. Пфлюгера. Позднее с помощью микроэлектродов был изучен механизм действия постоянного тока на ткани. Было установлено, что в области приложения к наружной поверхности мембраны, имеющей положительный заряд, положительного анода, происходит пассивная гиперполяризация и на этом участке возрастает положительный потенциал. При прикладывании отрицательного катода к наружной поверхности мембраны в этой области снижается положительный потенциал мембраны, так как происходит пассивной деполяризации. Изменения мембранного потенциала на участках прикладывания электродов к ткани называются пассивными, потому, что они происходят, не в результате изменения ионной проницаемости самой мембраны, а под влиянием катода (-) и анода (+). Пассивная гиперполяризация под анодом не сопровождается изменениями ионной проницаемости мембраны даже при большой силе приложенного тока. Поэтому, при замыкании постоянного тока возбуждение под анодом не возникает. В отличие от этого, пассивная деполяризация под катодом влечет за собой кратковременное повышение проницаемости мембраны для ионов Na+ и медленно развивающиеся, стойкое повышение проницаемости мембраны для ионов К+, поэтому, при замыкании постоянного тока возбуждение возникает под катодом. Таким образом, мгновенное замыкание приводит к постепенному возрастанию поляризации на аноде, т.е. гиперполяризации и уменьшению поляризации на катоде, т.е. деполяризации. При размыкании происходит медленное возвращение мембранного потенциала к исходному уровню. То, что замыкательный удар эффективнее размыкательного, так как порог раздражения при замыкании ниже, чем при размыкании, объясняется тем, что при наличии под катодом начальной деполяризации требуется очень незначительный раздражитель, чтобы вызвать дальнейшую деполяризацию под катодом, которая быстро достигает критического уровня деполяризации. Закон физиологического электротона. Согласно этому закону, при длительном прохождении постоянного тока в тканях изменяется их возбудимость. Она повышается на катоде - это называется катэлектротоном, а на аноде возбудимость снижается - это называется анэлектротоном. Эти изменения возбудимости, создаваемые прохождением постоянного тока носят название физиологического электротона (рисунок 2). Было также установлено, что при длительном прохождении постоянного тока в тканях возбудимость повышается не только непосредственно под катодом, но и на соседних с ним участках. Понижение возбудимости наблюдается не только под анодом, но и на соседних участках. Эти изменения возбудимости менее значительны по мере удаления от катода и анода, чем на участках, где непосредственно приложен анод и катод. В 1883 году Б.Ф. Вериго внес важное дополнение к закону Э.Ф. Пфлюгера: катод повышает возбудимость и возбуждает, а анод не возбуждает и понижает возбудимость. Он также показал, что при длительном действии сильного постоянного тока электротонические изменения возбудимости извращаются, т.е. под катодом начальное повышение возбудимости сменяется ее понижением и наступает катодическая депрессия, а под анодом сниженная возбудимость постепенно повышается и наступает анодическая экзальтация.
Рисунок 2. Физиологический электротон. Изменения мембранного потенциала и возбудимости при действии на возбудимую ткань постоянного тока а – изменения мембранного потенциала; б - изменения критического уровня деполяризации; в – изменения возбудимости Примечание: стрелкой отмечено начало действия тока.
История открытия биоэлектрических явлений. Первые исследования электрической активности живых тканей были проведены Л. Гальвани. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенной на медном крючке, при соприкосновении с железными перилами балкона (первый опыт Л. Гальвани). На основании этих наблюдений, им был сделан вывод, что сокращение лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам (крючку и перилам) к мышцам. Физик А. Вольта, повторив этот опыт, пришел к другому заключению. Источником тока, по его мнению, является не спинной мозг и «животное электричество», а разность потенциалов, образующаяся в месте контакта разнородных металлов – меди и железа, а нервно-мышечный препарат лягушки является лишь проводником электричества. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени, что вызывало сокращение мышцы (второй опыт Л.Гальвани), тем самым доказав существование «животного электричества». К. Маттеучи показал, что можно вызвать сокращение мышц нервно-мышечного препарата, прикладывая нерв к сокращающимся мышцам другого препарата. Этот опыт свидетельствует о том, что в сокращающейся мышце возникают токи настолько значительные, что их можно использовать в качестве раздражителя для нерва другого препарата. Эти токи получили название «токов действия». Позднее Э. Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва на поврежденный и неповрежденный участки мышцы, возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током повреждения. Мембранный потенциал. Происхождение и механизмы поддержания величины мембранного потенциала. В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая называется мембранным потенциалом (МП), или, если это клетка возбудимой ткани, - потенциалом покоя (рисунок 3).В состоянии покоя, внутренняя сторона мембраны заряжена отрицательно по отношению к наружной поверхности. Принимая потенциал наружного раствора за нуль, МП записывают со знаком «минус». Его величина у разных клеток колеблется от минус 30 до минус 100 мВ. Следовательно, в состоянии покоя клеточная мембрана находится в состоянии поляризации, уменьшение величины мембранного потенциала называется деполяризацией, увеличение – гиперполяризацией, восстановлениеисходного значения мембранного потенциала – реполяризацией мембраны.
I II
Рисунок 3.Схема регистрации мембранного потенциала I - графическое изображение мембранного потенциала II - схема регистрации мембранного потенциала
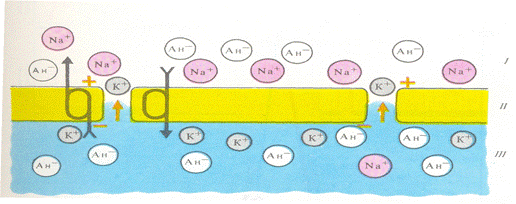
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном в1902 году. Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Г. Нернста. В 1949 – 1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали современную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Плазматическая мембрана практически не проницаема для внутриклеточных белков и других органических ионов. Ионы калия диффундируют из клетки по концентрационному градиенту, а непроникающие анионы остаются в цитоплазме, обеспечивая появление разности потенциалов через мембрану. Возникающая разность потенциалов препятствует выходу калия из клетки, при некотором ее значении наступает равновесие между выходом калия по концентрационному градиенту и выходом этих катионов по возникшему электрическому градиенту. Мембранный потенциал, при котором достигается это равновесие, называется равновесным потенциалом. Цитоплазма нервных и мышечных клеток содержит в 30 – 50 раз больше ионов калия, в 8 – 10 раз меньше ионов натрия, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами (рисунок 4), которые образованы макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, потенциалозависимые открываются и закрываются в ответ на изменения МП.
Рисунок 4. Ионные каналы клеточной плазматической мембраны Популярное:
|
Последнее изменение этой страницы: 2016-03-17; Просмотров: 3123; Нарушение авторского права страницы