|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ОСНОВНЫЕ ЯВЛЕНИЯ БИОЛОГИИ РАЗВИТИЯСтр 1 из 7Следующая ⇒
Основы Общей и сравнительной Эмбриологии
Киров 2008
УДК 611.013 ББК 28.03 0-75 Издается по решению ЦМС и РИС ГОУ ВПО «Кировская государственная медицинская академия» (протокол № от ) Основы общей и сравнительной эмбриологии: Учебное пособие для студентов медицинских вузов / Сост.: Е.В. Коледаева В.Б. Зайцев, И.Н.Гамулинская. – Киров: Кировская ГМА. - 2008. — 65 с: илл.?.
В пособии представлено подробное систематизированное изложение основ общей и сравнительной эмбриологии. Большое внимание уделено закономерностям эмбрионального развития птиц и млекопитающих. Подробно рассматриваются вопросы гистогенеза и органогенеза, необходимые для изучения студентами-медиками основ эмбриологии человека. Пособие снабжено значительным иллюстративным материалом и заданиями для самостоятельной работы студентов. Содержание пособия соответствует Программе по гистологии, цитологии и эмбриологии для высших медицинских учебных заведений (Москва, 2002). Эмбриологический словарь содержит необходимые термины для успешного усвоения учебного материла. Пособие предназначено для самостоятельной работы студентов медицинских Вузов. Рецензенты: Спицин А. П. – заведующий кафедрой патофизиологии Кировской государственной медицинской академии, доктор медицинских наук, профессор. Панфилов А.Б. — заведующий кафедрой гистологии, цитологии и эмбриологии, анатомии и патофизиологии Вятской государственной сельскохозяйственной академии, доктор ветеринарных наук, профессор.
© Е.В. Коледаева, В.Б. Зайцев, И.Н. Гамулинских, 2008 © ГОУ ВПО Кировская ГМА Росздрава Оглавление ПРЕДИСЛОВИЕ Эмбриология (от греч. Embrion – зародыш и Logos – учение) - это наука, изучающая закономерности развития зародыша до рождения (т.е. индивидуальное развитие, которое начинается оплодотворением и заканчивается рождением плода). Истоки эмбриологии теряются в веках. Становление эмбриологии как науки тесно связано с именем Карла Бэра, который сформулировал правило: развитие организма происходит от общего к частному; А.О. Ковалевского, который изучал сравнительную эмбриологию и выявил единый план развития ланцетника, червей, кишечнополостных; обосновал теорию зародышевых листков; Э. Геккеля – немецкого биолога – сформулировавшего биогенетический закон: «Онтогенез – есть краткое повторение филогенеза». Большой вклад в развитие эмбриологической науки внесли теории параллельного и дивергентного развития тканей, разработанные выдающимися учеными профессором А.А. Заварзиным и профессором М.Г. Хлопиным. Бурное развитие эмбриологии отмечается в последние несколько десятилетий. Оно обусловлено рядом выдающихся открытий в области генетики, иммунологии, молекулярной биологии, которые позволили переосмыслить некоторые этапы эмбрионального морфогенеза. Освоение эмбриологической науки играет огромную роль в подготовке современного врача, так как проблемы бесплодия, аномалий внутриутробного развития человека в настоящее время актуальны. Об этом свидетельствует действующая в нашей стране национальная программа по охране материнства и детства. Для освоения курса эмбриологии человека студент-медик должен иметь глубокие знания об эволюционном усложнении эмбрионального развития позвоночных. Данное учебное пособие предназначено для самостоятельного изучения общей и сравнительной эмбриологии для студентов лечебного и педиатрического факультетов медицинских вузов. Материал изложен в доступной форме с наглядными иллюстрациями, схемами и таблицами, что позволяет систематизировать изучаемый материал. Для закрепления знаний в пособии представлены ситуационные задачи и тестовые задания для студентов. Пособие содержит подробный современный словарь эмбриологических терминов. При подготовке учебного пособия использованы фундаментальные издания по эмбриологии: Кнорре А.Г. Краткий очерк эмбриологии человека. 1967г., Пэттен Б.М. Эмбриология человека. 1959г., Токин Б.П. Общая эмбриология. 1977г., а также издания современных эмбриологических школ, возглавляемых академиком РАМН Волковой О.В., профессором Вальковичем Э.И., профессором Даниловым Р.К.. Методические указания Учебное пособие предназначено для студентов лечебного и педиатрического факультетов и для аспирантов медицинских вузов, а также будет полезно для студентов факультетов биологической и ветеринарной медицины. Цель учебного пособия – дать подробное систематизированное изложение современных данных по общей и сравнительной эмбриологии. Учебное пособие включает задачи: 1) Рассмотреть основные явления биологии развития 2) Выявить отличия половых клеток от соматических 3) Изучить последовательные этапы собственно эмбриогенеза и их особенности 4) Изучить гистогенез и органогенез 5) Изучить эволюционную сравнительную характеристику эмбрионального развития, начиная от низших позвоночных до млекопитающих. Необходимыми для изучения темы смежными дисциплинами (базисные знания) являются: анатомия (гистофизиология половых органов, развитие мужских (сперматогенез) и женских (овогенез) половых клеток), биология (мейоз, типы яйцеклеток, бластул, гаструл низших позвоночных). Согласно учебной программе длительность изучения темы составляет 3 часа для лечебного факультета и 3 часа для педиатрического факультета, что составляет 3-4% общей учебной нагрузки. По данной теме читается лекция, разбор её осуществляется на практическом занятии. Самостоятельная работа студентов включает в себя посещения медицинской библиотеки, подготовку докладов к практическим занятиям, заседаниям научного кружка, а также решение ситуационных задач, контрольных тестов и диагностику гистологических микропрепаратов и электронограмм. В результате изучения данного раздела дисциплины студенты должны знать: классификацию яйцеклеток, типы дробления и гаструляции, комплекс осевых зачатков органов, дифференцировку зародышевых листков и особенности эмбрионального развития низших и высших позвоночных. Студенты должны уметь: диагностировать гистопрепараты бластулы, гаструлы и нейрулы лягушки, гистопрепараты зародыша курицы на стадии нейрулы и стадии образования провизорных органов, электронограммы сперматозоида и яйцеклетки, решать ситуационные задачи по теме.
ОСНОВНЫЕ ЯВЛЕНИЯ БИОЛОГИИ РАЗВИТИЯ Развитие организма представляет собой процесс систематического, последовательного, упорядоченного накопления структурных и функциональных качеств прогрессивного характера, происходящий сопряжено на всех уровнях структурной организации живой материи: молекулярном, клеточном, тканевом, органном, системо-органном и организменном. Эти изменения определены во времени и происходят в строгой последовательности, взаимосвязи и взаимообусловленности на всех уровнях структурной организации живого. Основными биологическими процессами развития являются: 1. Размножение клеток. 2. Рост. 3. Детерминация. 4. Дифференцировка. 5. Индукция. 6. Интеграция. 7. Апоптоз. Размножение клеток. Деление клеток имеет важнейшее значение для многих процессов развития, представляя собой одно из основных свойств живых систем. Следствием деления является увеличение числа клеток, являющееся основным механизмом роста, как вэмбриональный, так и в постэмбриональный периоды развития. Для формирования конкретных структур зародыша требуется определенное минимальное число клеток. Рост. Если сравнить массу человеческой яйцеклетки диаметром 0, 15 мм с массой тела взрослого человека, то становится очевидным, что в процессе развития происходит рост поистине астрономических масштабов. Наиболее просто рост можно определить как увеличение массы ткани органа за счет увеличения числа клеток, т. е. гиперплазии. Увеличение массы всегда сопровождается увеличением линейных размеров. В то же время масса может возрастать за счет увеличения размеров клеток, т. е. гипертрофии. Рост регулируется гормоном роста и некоторыми химическими веществами, например, фактором роста фибробластов, эритропоэтином, кейлонами и др. Дифференциальный рост — это различная скорость роста одних и тех же тканей в разных участках тела и в разное время. Аллометрический рост — это диспропорциональный рост отдельных частей тела в постэмбриональный период. Детерминация. Детерминация (от латинского determinare — определять) — процесс определения пути развития материала эмбриональных зачатков в направлении образования специфических тканевых структур. Детерминация клеток обусловлена процессами репрессии и дерепрессии генов, происходящими на уровне транскрипции кода. В процессе детерминации, по мере появления признаков специализации, потенции клеток зародыша к развитию в иных направлениях суживаются. В этих случаях принято говорить о процессе рестрикции (ограничении). Детерминация может быть оотипической, зачатковой и тканевой в зависимости от этапов развития клеток зародыша. а) Оотипическая. Реализуется на стадии яйцеклетки и зиготы. На данном этапе существуют биохимические различия не между отдельными клетками, а между различными участками цитоплазмы зиготы; эти участки называются презумптивными. Попадая при дроблении в тот или иной бластомер, каждый такой участок определяет направление дальнейшей дифференцировки этого бластомера. б) Бластомерная(зачатковая). Здесь уже имеются биохимические и морфологические различия между бластомерами, которые усиливаются по мере дробления. До стадии 4—8 бластомеров различия еще не очень велики (почему возможно появление однояйцевых близнецов), а затем они становятся необратимыми. в) Тканевая. Дальнейшие этапы образования зародышевых листков, появления и формирования тканей. При этом различаются лабильная и стабильная детерминация. Лабильная детерминация еще предполагает возможность некоторой изменчивости свойств клеток под влиянием условий внешней среды, в частности, при экспериментальных исследованиях. Стабильная детерминация характеризуется стойким закреплением свойств клеток, способных синтезировать специфические белки. Н. Г. Хлопин определил гистологическую детерминацию как тканевую наследственность, проявляющуюся в высокой специфичности всех свойств ткани, сохраняющихся в любых нормальных и патологических условиях. Дифференцировка. Дифференцировка (от латинского differentia — различие) — это развитие клеток, проявляющееся в их изменении из ранее однородных (индифферентных) в неоднородные, различным образом специализированные клетки. Дифференцировка клеток протекает в период интерфазы и представляет собой результат реализации в данных условиях генетической информации, исходящей из ДНК клеточного ядра, и проявляется в синтезе специфических белков. С морфологической точки зрения вследствие дифференцировки в развивающихся клетках появляются специальные органеллы, выполняющие определенные
специфические функции (например: миофибриллы, нейрофибриллы, тонофибриллы, реснички и т. д.). По мнению А. Г. Кнорре (1971), необходимо различать: 1. Дифференцировку органологическую, или пространственную, т. е. анатомическое обособление частей организма или органа. 2. Дифференцировку гистологическую, т. е. появление местных различий в клеточном материале различных участков зародыша, приводящее к образованию тканей. 3. Дифференцировку клеточную, в результате которой в первоначально однородной цитоплазме появляются разнородные структуры, а клетка приобретает специфическую форму. Таким образом, дифференцировка связана со специализацией клеток, их комплексов, тканей и органов, направленной на выполнение определенных, свойственных этим частям организма, функций. Клеточная дифференцировка протекает на всех этапах онтогенеза: дозачатковом, зачатковом и тканевом. Индукция. Индукция (от латинского inductio — наведение, побуждение) — взаимовлияние, взаимодействие одних клеток или эмбриональных зачатков (индукторов) на другие клетки или зачатки, в результате чего направление развития последних становится качественно иным по сравнению с тем, каким оно бы было в отсутствие действия индуктора. Так, при взаимодействии хордомезодермы с эктодермой из последней развивается нервная система; глазной пузырь, контактируя с эктодермой, индуцирует развитие хрусталика глаза и т. д. Индукция — это процесс дифференцировки, вызванный действием двух зачатков друг на друга, происходящим благодаря выработке морфогенетических сигналов и их передаче соответствующему рецептору. Интеграция. Интеграция (от латинского integratio — объединение) — объединение клеток в систему, установление между ними взаимосвязи и взаимообусловленности в процессе их развития. Интеграция означает взаимосвязь и взаимозависимость, затрагивая все сферы жизнедеятельности клеток: обмен веществ, рост, дифференцировку, специализацию, размножение, инволюцию, клеточную гибель и др. К интегрирующим факторам относятся сосудистые, гуморальные, нервные, эндокринные, иммунные, способные оказывать существенное влияние на ход гисто- и органогенеза, обеспечивая целостность организма в онтогенезе. Апоптоз Апоптоз (клеточная гибель) — очевидный компонент многих стадий развития, необратимо генетически детерминированный процесс, играющий важную роль в развитии зародыша. Созревание. Конечный этап развития (дифференцировки и роста), необходимый для подготовки тканей и органов или систем органов к выполнению специальной для них функции, называется созреванием. Морфогенез. Группа процессов, формирующих внешнюю и внутреннюю конфигурацию зародыша, объединяется под общим названием морфогенез, который делится на две основные фазы: 1) гистогенез — развитие из материала эмбриональных зачатков дефинитивных тканей; 2) органогенез — образование и развитие органов зародыша. Обычно органогенез протекает одновременно с гистогенезом, хотя между этими процессами отмечается относительная независимость.
I. Прогенез Период развития половых клеток, предшествующий началу собственно индивидуального развития организма, называется гаметогенезом. Этот процесс осуществляется в ходе этапа развития, известного под названием прогенез. I.1 Гаметогенез Гаметогенез — это процесс образования и созревания половых клеток в половых железах; мужских половых клеток — сперматозоидов (сперматогенез) и женских гамет — яйцеклеток (овогенез). Мейоз — деление половых клеток, результатом которого является образование гамет с гаплоидным набором хромосом. Кроме того, во время мейоза создаются новые комбинации генетического материала родителей, которые достигаются в результате: — случайного распределения материнских и отцовских хромосом по дочерним клеткам; — обмена участками гомологичных хромосом при кроссинговере. Мейоз состоит из двух следующих одно за другим делений клетки: редукционного и эквационного. К началу профазы первого деления в стадию прелептотены в половой клетке завершается удвоение ДНК, хотя количество хромосом не изменяется (Рис. 1). Профаза первого деления (редукционного) состоит из 5 стадий: лептотены, зиготены, пахитены, диплотены, диакинеза. Лептотена. Хромосомы имеют форму тонких, длинных нитей, закручивающихся в спираль. Они состоят из двух идентичных сестринских хроматид, которые соединены в одном месте общей центромерой. Различить отдельные хроматиды с помощью обычного светового микроскопа невозможно. Зиготена. Гомологичные парные хромосомы сближаются и располагаются одна подле другой, образуя биваленты (конъюгация). Область контакта между конъюгированными хромосомами имеет особую структуру и называется синаптонемальным комплексом(СК), принимающим участие в кроссинговере. Синаптонемальный комплекс представляет собой лентообразную структуру. На периферии СК расположены более темные полосы – осевые тяжи хромосом, а в центре – более светлая центральная зона, по середине которой идет темная центральная полоса. От последней отходят к осевым тяжам многочисленные тонкие филаменты, создающие мелкую поперечную исчерченность СК. СК имеет белково-полисахаридную природу и обеспечивает точность кроссинговера.
Пахитена . Гомологичные хромосомы обмениваются генетическим материалом. Этот процесс называется кроссинговером. Конъюгация заканчивается, происходит утолщение хромосом, обусловленное спирализацией, оставаясь в составе бивалентов. Диплотена. Отдельные участки конъюгировавших хромосом перекрещиваются друг с другом. Те места, в которых они при этом соприкасаются, называются хиазмами, и считается, что именно здесь гомологичные материнские и отцовские хромосомные нити разрываются, и происходит обмен участками в процессе кроссинговера. На стадии диплотены происходит разделение двух конъюгировавших хромосом в результате продольного расщепления по всей длине, за исключением хиазм синаптонемального комплекса. Каждая пара хромосом выглядит теперь как 4 попарно связанные хроматиды и называется тетрадой. Диакинез. Хромосомы еще более укорачиваются, расщепление их на хроматиды делается более заметным, хиазмы перемещаются к концам хромосом. В это время исчезают ядрышко и ядерная оболочка, становится заметным аппарат веретена деления. В метафазупервого деления тетрады выстраиваются так, что если материнская хромосома (диада) оказывается по одну сторону плоскости экватора, то отцовская — по другую. Распределение же отцовских и материнских хромосом по ту или иную сторону экватора происходит случайным образом. В анафазу происходит перемещение хромосом (диад) к полюсам клетки. В отличие от обычного митоза не происходит разрывов в области центромеров хромосом. К одному полюсу отходит целая материнская хромосома, к другому — отцовская. Это приводит к образованию генетически неоднородных дочерних клеток. В то же время при расхождении хромосом происходит разрыв хиазм и, таким образом, завершается кроссинговер. Далее происходит обычная телофаза . Поскольку каждая хромосома по-прежнему образована двумя хроматидами, хромосомный набор клеток, образовавшихся в результате первого деления мейоза, может быть представлен формулой 1n (гаплоидное число хромосом) 2с (каждая хромосома содержит 2 сестринские хроматиды). Особенностью мейоза является отсутствие периода синтеза (не происходит репликация ДНК) между первым и вторым делениями мейоза. Второе деление мейоза (эквационное) протекает подобно обычному митотическому с той лишь разницей, что в деление вступают клетки, не реплицировавшие ДНК (1n2с). Вследствие этого образуются гаметы с гаплоидным набором хромосом. Мужские половые клетки – сперматозоиды – образуются в процессе сперматогенеза в мужских половых гонадах. Женские половые клетки – яйцеклетки – образуются в процессе овогенеза в женских половых гонадах. Половые клетки имеют ряд существенных отличий от соматических: 1. Гаплоидный набор хромосом: мужские - содержат или X, или Y хромосомы; женские - содержат только X хромосомы. 2. Невысокий уровень метаболизма. 3. Неспособность к размножению. 4. Специфические ядерно-цитоплазматические отношения: У сперматозоида смещено в сторону ядра, т.к. клетка специализирована на несении наследственной информации; У яйцеклетки - в сторону цитоплазмы, т.к. клетка выполняет трофическую функцию. 5. Закладка половых клеток происходит вне тела эмбриона, в энтодерме стенки желточного мешка.
I.3 Морфология яйцеклетки.
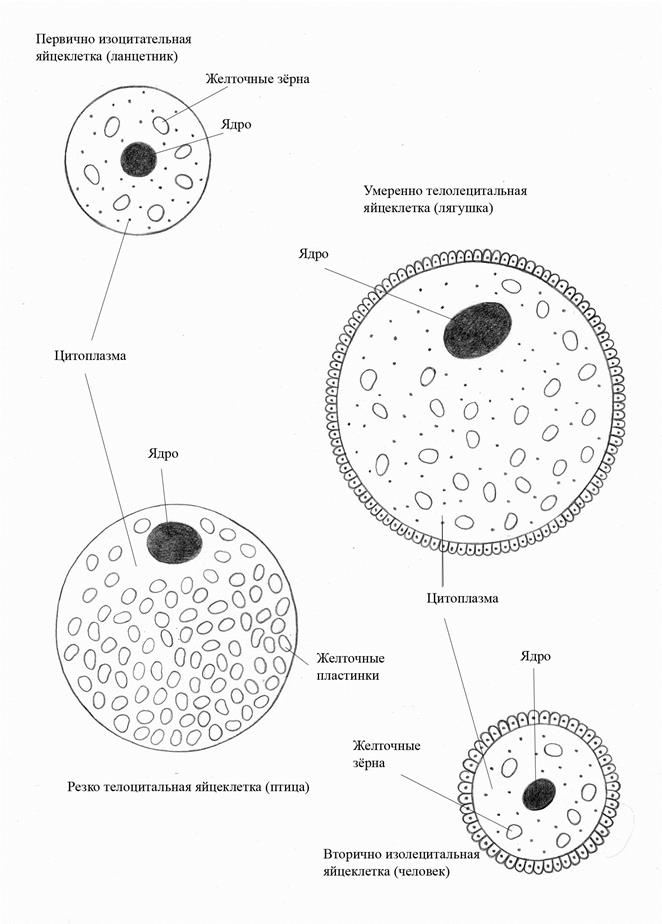
Яйцеклетки разных животных имеют сходные черты: · округлая форма, крупная; · диаметр 130-140 мкм; · неподвижная. Ядро с преобладанием эухроматина в цитоплазме распологаются митохондрии, аппарат Гольджи, гранулярная ЭПС, отсутствуют центриоли. Содержит желточные гранулы, которые отграничены мембраной и содержат фосфовитин и липовителлин. Эти вещества имеют как энтогенное, так и экзогенное происхождение: часть из них образуется в овоците, часть – в печени. Также в цитоплазме обнаруживаются мультивезикулярные тельца, представляющие собой совокупность мелких пузырьков, заключенных в большой мембранный мешок. Они представляют собой производные лизосом. Ещё один вид производных лизосом – кортикальные гранулы, которые содержат гидролитические ферменты и гликозаминогликаны (ГАГ), участвующие в развитии кортикальной реакции. На поверхности плазмолеммы выявляются многочисленные микроворсинки. Вокруг яйцеклетки обычно имеется несколько оболочек: 1. блестящая оболочка (10 мкм) – состоит из 3 фракций гликопротеидов: ZP1, ZP2, ZP3. С фракцией ZP3 связывают способность распознавать и связывать сперматозоид. 2. лучистый венец (corona radiata) состоящий из фолликулярных клеток. Они выполняют трофическую функцию и создают микроокружение для развивающегося овоцита. 3. в связи с наземным существованием пресмыкающихся и птиц возникла необходимость в третьей оболочке – скорлуповой (для предохранения от повреждающих факторов окружающей среды).
II Собственно эмбриогенез II.1 ОПЛОДОТВОРЕНИЕ Оплодотворение – это процесс слияния мужских и женских половых клеток, в результате чего восстанавливается диплоидный набор хромосом и происходит образование зиготы, которая является одноклеточным зародышем. Слияние гамет может произойти только при условии совместного действия многих биологических факторов. Циклы развития половых клеток должны протекать синхронно так, чтобы сперматозоиды и яйцеклетки созревали и выделялись в одни и те же сроки. Оплодотворению предшествует осеменение — процесс, приводящий к контакту мужской и женской половых клеток. В оплодотворении различают три фазы: 1) сближение и дистантное взаимодействие гамет, 2) контактное взаимодействие гамет, 3) проникновение сперматозоида (точнее, его ядра и центриолей) в яйцеклетку. После этого происходит еще одна важная стадия — четвертая — подготовка зиготы к дроблению. II.2 Дробление. Дробление представляет собой серию митотических делений зиготы с образованием многих дочерних клеток (бластомеров) меньшего размера. Отличием дробления от митоза является то, что бластомеры не растут и не расходятся. Эти процессы сдерживает оболочка оплодотворения, которая сохраняется вплоть до гаструляции. II.2.1 Типы дробления. Типы дробления зависят от типа яйцеклетки. Различают (Рис. 3): · полное (голобластическое) и неполное (меробластическое); · синхронное и асинхронное (клетки делятся не одновременно); · равномерное и неравномерное (образуются бластомеры разного размера). В результате дробления образуется бластула, содержащая полость – бластоцель. II.2.2 Типы бластул. Различают четыре типа дробления и столько же типов бластул (Рис. 4). У ланцетника дробление полное равномерное синхронное, в результате которого образовалась – целобластула, имеющая бластоцель, расположенную по середине и однослойную бластодерму. У амфибии – дробление полное неравномерное асинхронное, в результате образовалась – амфибластула, имеющая многослойную бластодерму и бластоцель, расположенную эксцентрично у анимального полюса. У рептилий и птиц – дробление неполное неравномерное асинхронное (дискоидальное). В результате образуется дискобластула, которая включает три компонента: · плоский зародышевый диск, образующийся на апикальной части и состоящий из небольших клеток, · нераздробившийся желток, · бластоцель, узкую щель между диском и желтком. У млекопитающих и человека дробление полное неравномерное асинхронное. В результате формируется бластоциста. Бластоциста состоит из трех компонентов: 1. Трофобласт – однослойная стенка, состоящая из светлых мелких бластомеров, из которой затем развивается внезародышевый орган – хорион. 2. Эмбриобласт – скопление крупных темных бластомеров на внутренней
Рисунок 4 Типы бластул
поверхности трофобласта у одного из полюсов. Источник развития самого зародыша и остальных внезародышевых органов (амнион, желточный мешок и аллантоис). 3. Бластоцель. На стадии бластулы окончательно устанавливается полярность зародыша. На стадии поздней бластулы устанавливаются так называемые презумптивные области (предположение, основанное на вероятности), которые создают материал определенных органов и систем. С помощью метода маркировки участков зародыша витальными красителями (метод В. Фогта, 1925 г.) были получены карты презумптивных зачатков. II.3 Гаструляция Направленные клеточные движения происходят на стадии гаструляции. Гаструляция (от лат. gaster — желудок) — сложный процесс химических и морфогенетических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки: наружный (эктодерма), средний (мезодерма) и внутренний (энтодерма) —источники зачатков тканей и органов. Способы гаструляции: 1. деляминация (расщепление), 2. инвагинация (впячивание), 3. иммиграция (выселение), 4. эпиболия (обрастание). В результате гаструляции возникает зародыш – гаструла. Гаструла имеет полость - гастроцель (полость первичной кишки), в которую ведет отверстие - бластопор (первичный рот). В зависимости от дальнейшей судьбы бластопора все животные подразделяются на: • первичноротых (беспозвоночные) - ротовое отверстие образуется на месте бластопора; • вторичноротых (хордовые и некоторые беспозвоночные) - бластопор преобразуется в анальное отверстие, а ротовое отверстие прорывается на брюшной стороне тела. У бластопора различают губы: дорсальную, латеральные и вентральную. Итогом гаструляции является формирование так называемого осевого комплекса зачатков: III.1.1. Тип яйцеклетки. Яйцеклетка относится к первичному изолецитальному типу (Рис. 2). Желтка в яйцеклетке немного, желточные гранулы распределены равномерно лишь с небольшим преобладанием в вегетативном полушарии по сравнению с анимальным. Анимальный полюс яйцеклетки приблизительно соответствует будущему переднему концу тела зародыша, т. е. еще до оплодотворения возникает переднезадняя ось тела. Сперматозоид проникает в яйцеклетку в одной из точек несколько ниже экватора. III.1.2. Стадия зиготы. На поверхности зиготы в области вхождения сперматозоида возникает так называемый зернистый серп, как его зеркальное отражение формируется серый серп. Последний отличается слабой пигментацией. Зернистый серп представляет собой область, где концентрируются митохондрии. У зародыша ланцетника на стадии зиготы уже выявляются презумптивные области, содержащие материал будущих зачатков: в анимальном полушарии зиготы содержится материал будущей эктодермы, в вегетативном полушарии — материал энтодермы, область серого серпа содержит материал двух зачатков — нервной пластинки (граничит с эктодермой) и хорды (граничит с энтодермой), в области зернистого серпа располагается материал мезодермы. Серый серп определяет будущую дорсальную поверхность тела зародыша, зернистый серп — вентральную поверхность. Через середины серпов проходит плоскость билатеральной симметрии. III.1.3.Тип дробления. Дробление у ланцетника полное равномерное синхронное (Рис.3). Зигота делится на бластомеры примерно одинаковой величины (вегетативные бластомеры чуть крупнее анимальных, так как в вегетативном полушарии содержалось несколько больше желтка). Первая борозда дробления — меридиональная — возникает у анимального полюса и, распространяясь к вегетативному полюсу, разделяет зиготу на два бластомера. При этом плоскость первого деления дробления проходит через середины серого и зернистого серпов, в результате чего образующиеся бластомеры по содержащемуся в них материалу идентичны зиготе. Если на этой стадии разделить бластомеры, каждый из них будет развиваться в самостоятельный организм. Вторая борозда также меридиональная, но проходит в плоскости, перпендикулярной по отношению к первой. Третья борозда, называемая экваториальной (или широтной), проходит несколько вышеэкватора.Затем происходит чередование меридиональных и широтных борозд. Бластомеры делятся синхронно, так что при каждом делении число бластомеров увеличивается вдвое, нарастая в геометрической прогрессии, которая нарушается лишь к концу дробления. III.1.5. Тип гаструляции. У ланцетника гаструляция происходит путем простого впячивания — инвагинации — вегетативной части бластулы внутрь бластоцели. У образующегося двухслойного зародыша — гаструлы — наружный листок представляет собой первичную эктодерму, содержащую материал эктодермы и нервной пластинки, внутренний листок является первичной
энтодермой, в которую помимо энтодермы входит материал мезодермы и хорды (Рис. 5). Гаструла по мере роста начинает плавать на одной из сторон, которая становится брюшной (вентральной), а противоположная ей сторона — спинной (дорсальной). У ланцетника, как и у всех хордовых животных, на месте бластопора образуется анальное отверстие, а дефинитивный рот возникает на противоположном конце. Край бластопора, связанный с материалом нервной пластинки и хорды, является дорсальным краем, или губой. Противоположный край бластопора называют вентральной губой, он связан с материалом мезодермы. Различают также боковые, или латеральные, губы бластопора, содержащие материал мезодермы. Во время гаструляции, в отличие от дробления, происходит активное перемещение клеточного материала. Растут и удлиняются зачатки нервной трубки и хорды. Материал мезодермы из области боковых и вентральной губ бластопора перемещается на дорсальную сторону тела зародыша. После того как весь материал мезодермы переместится на дорсальную сторону, у зародыша возникает осевой комплекс зачатков, типичный для всех хордовых животных. III.2.1.Тип яйцеклетки. Яйцеклетки низших позвоночных богаты желтком, относятся к умереннотелолецитальному типу (Рис. 2) и имеют более крупные, чем у ланцетника, размеры в связи с увеличением размеров тела. Яйцеклетка амфибий обладает полярностью, обусловленной неравномерным распределением желтка, и уже на этой стадии определяется переднезадняя ось тела зародыша. Анимальный полюс примерно соответствует будущему переднему концу тела зародыша, вегетативный полюс — заднему. III.2.2. Стадия зиготы. Сперматозоид проникает в яйцеклетку несколько ниже ее экватора. У амфибий на поверхности оплодотворенного яйца против места вхождения сперматозоида возникает слабо пигментированная серповидная зона — серый серп. Область серого серпа отмечает будущую дорсальную сторону тела зародыша, противоположная сторона соответствует будущей вентральной стороне тела. Место вхождения сперматозоида определяет дорсо-вентральную ось, а через середину серого серпа проходит плоскость билатеральной симметрии. III.2.3.Тип дробления. Дробление полное неравномерное асинхронное (Рис. 3). Первые две борозды дробления — меридиональные, проходят так же, как и у ланцетника, но распространяются от анимального полюса к вегетативному очень медленно в связи с загруженностью вегетативного полушария желтком. Третья борозда дробления — широтная, проходит значительно выше экватора, за счет чего уже на стадии восьми бластомеров появляются резкие различия в величине между анимальными и вегетативными бластомерами. Анимальные бластомеры мелкие, так как делятся значительно быстрее вегетативных. Кроме меридиональных и широтных борозд появляются тангенциальные борозды, проходящие в плоскости, параллельной поверхности зародыша, в результате чего стенка бластулы становится многослойной. III. 2.5. Тип гаструляции. У амфибий – два способа гаструляции: инвагинация происходит лишь в незначительной степени, и основным способом гаструляции является эпиболия (обрастание). Мелкие анимальные бластомеры за счет быстрого их деления начинают нарастать на более крупные вегетативные и почти полностью их окружают. За счет обрастания и, частично, впячивания бластомеры, содержащие материал энтодермы, прехордальной пластинки и хорды, оказываются внутри зародыша, образуя его внутренний листок. Бластомеры, содержащие материал нервной пластинки и эктодермы, располагаются снаружи, составляя наружный зародышевый листок, а клетки презумптивной мезодермы, находившейся на границе между двумя листками, начинают вклиниваться (иммиграция) в пространство между ними, продвигаясь на дорсальную сторону и к переднему концу зародыша, формируя средний зародышевый листок (Рис. 5). Таким образом, у амфибий к процессам эпиболии и инвагинации прибавляется иммиграция. После перемещения материала мезодермы на дорсальную сторону она принимает вид двух продольных тяжей, лежащих по бокам от зачатка хорды, латерально она врастает в пространство между наружным и внутренним листками, смыкаясь на вентральной стороне. У амфибий бластопор имеет кольцевидную форму (кольцевидный бластопор). Его просвет постоянно закрыт выступающей через него энтодермой (эту часть энтодермы называют «желточной пробкой»). У амфибий и других позвоночных, в отличие от ланцетника, в результате гаструляции образуется три зародышевых листка. К концу гаструляции одновременно с зародышевыми листками возникает осевой комплекс зачатков. IV.3.1.Тип яйцеклетки. Популярное:
|
Последнее изменение этой страницы: 2016-03-15; Просмотров: 1620; Нарушение авторского права страницы