|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Анатомо-морфологическая база высших психических функций.
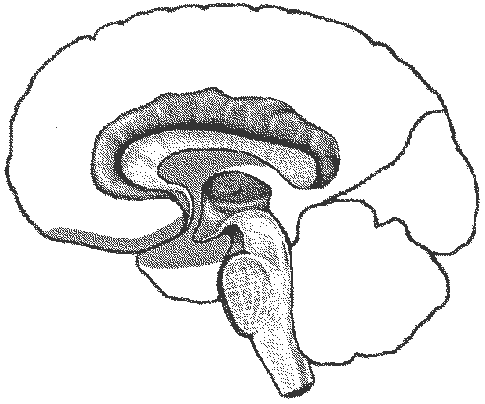
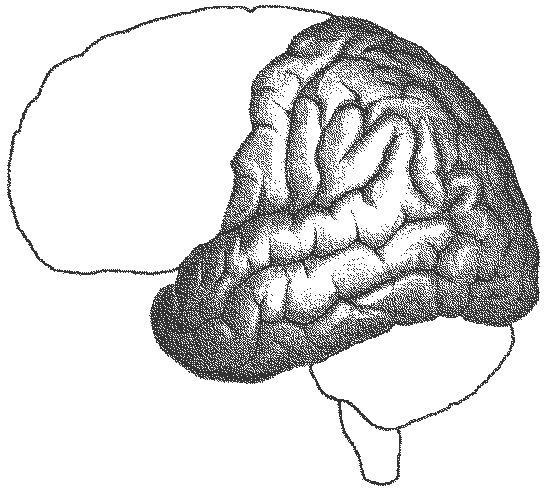
Мозг человека как специальный орган, осуществляющий высшую форму обработки информации, представляет лишь часть нервного аппарата — системы, специализирующейся на согласовании внутренних потребностей организма с возможностями их реализации во внешней, в том числе социальной, среде. Как и всякая система, она имеет определенную пространственную и функциональную конструкцию, сформировавшуюся в ходе эволюционного процесса. Поэтому диапазон основных параметров функционирования нервной системы в целом отражает вероятностную структуру качества и интенсивности раздражителей, с которыми формирующийся организм сталкивался на протяжении фило- и онтогенеза. Нервная система с входящим в нее мозгом — это иерархически и функционально упорядоченное материальное пространство, являющееся неотъемлемым элементом более общей системы — организма. Наиболее дифференцированным отделом ЦНС является кора головного мозга, которая по морфологическому строению в основном делится на шесть слоев, отличающихся по строению и расположению нервных элементов. Прямые физиологические исследования коры доказали, что ее основной структурно-организующей единицей являетсятак называемая кортикальная колонка, представляющая собой вертикальный нейронный модуль, все клетки которого имеют общее рецепторное поле или однородно функционально ориентированы. Колонки группируются в более сложные образования — макроколонки, сохраняют определенный топологический порядок и образуют строго связанные распределенные системы. Благодаря исследованиям Бродмана, О. Фогта и Ц. Фогт и работам сотрудников Московского института мозга было выявлено более 50 различных участков коры — корковых цитоархитектонических полей, в которых нервные элементы имеют свою морфологическую и функциональную специфику. [См. Хомская Е. Д. Нейропсихология. — М., 1987.] Кора головного мозга, подкорковые структуры, а также периферические компоненты организма связаны волокнами нейронов, образующими несколько типов проводящих путей, связывающих между собой и различные отделы ЦНС. Существует несколько способов классификации этих путей, наиболее общий из которых предусматривает пять вариантов. Существенным смысловым компонентом подобной схемы являетсятезис, в соответствии с которым различные типы волокон являются представителями различных систем мозга, обеспечивающими разнообразный психофизиологический эффект их работы. Ассоциативные волокна — проходят внутри только одного полушария и связывают соседние извилины в виде коротких дугообразных пучков, либо кору различных долей, что требует более длинных волокон. Назначение ассоциативных связей — обеспечение целостной работы одного полушария как анализатора и синтезатора разномодальных возбуждений. Проекционные волокна — связывают периферические рецепторы с корой головного мозга. С момента входа в спинной мозг это восходящие афферентные пути, имеющие перекрест на различных его уровнях или на уровне продолговатого мозга. Их задача — трансляция мономодального импульса к соответствующим корковым представительствам того или иного анализатора. Почти все проекционные волокна проходят через таламус. Интегративно-пусковые волокна — начинаются от двигательных зон мозга, являются нисходящими эфферентными и по аналогии с проекционными также имеют перекресты на различных уровнях стволового участка или спинного мозга. Задача этих волокон — синтез возбуждений разной модальности в мотивационно организованную двигательную активность. Окончательной зоной приложения интегративно-пусковых волокон является мышечный аппарат человека. С точки зрения их топологической организации они также могут рассматриваться и как проекционные, поскольку реализуют принцип строгого соответствия (фактически — связи) между центральными корковыми нейронными группами и периферическими мышечными волокнами. Комиссуральные волокна — обеспечивают целостную совместную работу двух полушарий. Они представлены одним крупным анатомическим образованием — мозолистым телом, а также несколькими более мелкими структурами, важнейшими из которых являются четверохолмие, зрительная хиазма и межуточная масса таламуса. Функционально мозолистое тело состоит из трех отделов: переднего, среднего и заднего. Передний отдел обслуживает процессы взаимодействия в двигательной сфере, средний — в слуховой и слухоречевой, а задний — в тактильной и зрительной. Предположительно большая часть волокон мозолистого тела участвует в межполушарных ассоциативных процессах, регуляция которых может сводиться как к взаимной активации объединяемых участков мозга, так и к торможению деятельности контралатеральных зон. Лимбико-ретикулярные волокна — связывают энергорегулирующие зоны продолговатого мозга с корой. Задача этих путей — поддержание циклов общего активного или пассивного фона, выражающихся для человека в феноменах бодрствования, ясного сознания или сна. Область распространения ретикулярной формации точно не установлена. На основании физиологических данных, она занимает центральное положение в продолговатом мозге, мосте, среднем мозге, в гипоталамической области и даже в медиальной части зрительных бугров. Наиболее мощные связи продолговатый мозг образует с лобными долями. Определенная часть ретикулярных волокон обслуживает и работу спинного мозга. Морфогенез мозга определяется размерами и различием по клеточному составу как целого мозга, так и его отдельных структур. Кроме того, полноценный анализ зрелого мозга предусматривает и оценку характера взаимосвязи и способа организации различных частей мозга — нейронных ансамблей (Корсакова, Микадзе, Балашова). Масса мозга как общий показатель изменения нервной ткани составляет при рождении примерно (данные различных авторов колеблются) 390 г у мальчиков и 355 г у девочек и увеличивается соответственно до 1353 и 1230 г к моменту полового созревания. Наибольшее увеличение мозга происходит на первом году жизни и замедляется к 7-8 годам, достигая максимальной массы (примерно 1400 г) у мужчин к 19—20, а у женщин — к 16-18 годам. При рождении у ребенка полностью сформированы подкорковые образования и те области мозга, в которых заканчиваются нервные волокна, идущие от периферических частей анализаторов. Остальные зоны еще не достигают необходимого уровня зрелости, что проявляется в малом размере входящих в них клеток, недостаточном развитии ширины их верхних слоев, выполняющих в дальнейшем самую сложную ассоциативную функцию, незавершенностью в развитии проводящих нервных волокон. Скорость роста коры во всех областях мозга в целом наиболее высока в первый год жизни ребенка, но в разных зонах она заметно отличается. К 3 годам происходит замедление роста коры в первичных отделах, а к 7 годам — в ассоциативных. У трехлетних детей клетки коры уже значительно дифференцированы, а у 8-летнего мало отличаются от клеток взрослого человека. По некоторым данным от рождения до 2 лет происходит активное образование контактов между нервными клетками (через синапсы) и их количество в этот период выше, чем у взрослого человека. К 7 годам их число уменьшается до уровня, свойственного взрослому. Более высокая синаптическая плотность в раннем возрасте рассматривается как основа усвоения опыта. Исследования показали, что процесс миелинизации, по завершению которого нервные элементы готовы к полноценному функционированию, в разных частях мозга также проходит неравномерно. В первичных зонах анализаторов он завершается достаточно рано, а в ассоциативных — затягивается на длительный срок. Миелинизация двигательных корешков и зрительного тракта завершается в первый год после рождения, пирамидного тракта, задней центральной извилины (в которой осуществляется проекция кожной и мышечно-суставной чувствительности) — в 2 года, передней центральной извилины (начала двигательных путей) — в 3 года, слуховых путей — в 4 года, ретикулярной формации (энерго- и ритморегулирующей системы) — в 18 лет, ассоциативных путей — в 25 лет. Формирование большинства функциональных мозговых структур, относительно надежно способных реализовывать ту или иную психическую или психофизиологическую функцию в меняющихся условиях среды — нейронных ансамблей, заканчивается в 18 лет, кроме лобной области, где этот процесс завершается к 20 годам, а в префронтальных участках, по некоторым данным, и позднее. С точки зрения функциональных возможностей мозга раньше всех в эмбриогенезе закладываются предпосылки для становления кожно-кинестетического и двигательного анализаторов. В кожно-кинестетическом анализаторе первые два года — это этап формирования целевых специализированных действий. Способность к тонкому анализу проприоцептивных (кинестетических) раздражений появляется с 2-3 месяцев и развивается до 18-20 лет. Слуховые рецепторы начинают функционировать сразу после рождения, а на стыке 1 и 2 лет происходит усиленное образование условных рефлексов на речь. Тонкая дифференцировка звуковых раздражителей продолжается до 6-7 лет. Анализ вызванных потенциалов в корковых полях, вовлекаемых в зрительное восприятие, показывает, что специализация полей в первые 3-4 года невелика. В дальнейшем она нарастает и достигает наибольшей выраженности к 6-7 годам. Это позволяет рассматривать возраст 6-7 лет как сенситивный в становлении системной организации зрения (условные рефлексы со слухового анализатора начинают вырабатываться раньше, чем со зрительного). Ассоциативные отделы мозга прогрессируют поэтапно — «пик» первого этапа примерно совпадает с 2 годами, а второго — с 6-7 годами. Наиболее медленным темпом развития характеризуются, как уже указывалось, лобные отделы мозга, функцией которых является произвольная (в том числе и опосредованная речью) регуляция всех видов психической деятельности. Функциональные блоки мозга. На основе изучения нарушений психических процессов при различных локальных поражениях центральной нервной системы Лурия разработал общую структурно-функциональную модель мозга как субстрата психики. Согласно этой модели весь мозг может быть разделен на три основных блока, характеризующихся определенными особенностями строения и ролью в исполнении психических функций. 1-й блок — энергетический — включает ретикулярную формацию ствола мозга, неспецифические структуры среднего мозга, диэнцефальные отделы, лимбическую систему, медиобазальные отделы коры лобных и височных долей (рис. 16).
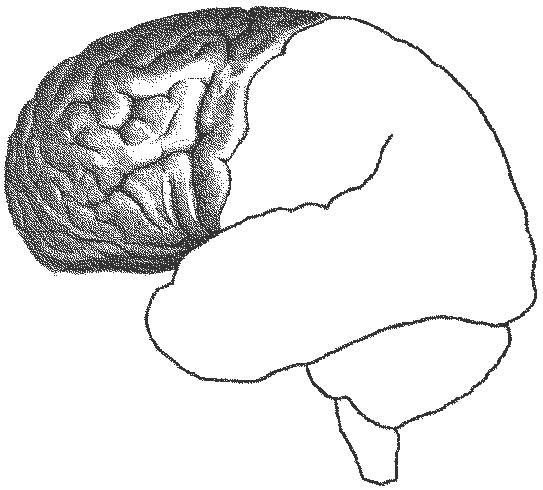
Рис. 16. Функциональные блоки мозга — 1-й блок (по Лурия). Блок регулирует общие изменения активации мозга (тонус мозга, необходимый для выполнения любой психической деятельности, уровень бодрствования) и локальные избирательные активационные изменения, необходимые для осуществления ВПФ. При этом за первый класс активаций несет ответственность преимущественно ретикулярная формация ствола мозга, а за второй — более высоко расположенные отделы — неспецифические образования диэнцефального мозга, а также лимбические и корковые медиобазальные структуры. Ретикулярная формация (РФ) обнаружена в 1946 г. в результате исследований американского нейрофизиолога Мегоуна, который показал, что эта клеточная функциональная система имеет отношение к регуляции вегетативной и соматической рефлекторной деятельности. Позднее совместными работами с итальянским нейрофизиологом Моруцци было продемонстрировано, что раздражение ретикулярной формации эффективно влияет и на функции высших структур мозга, в частности коры больших полушарий, определяя ее переход в активное, бодрствующее или в сонное состояние. Исследования показали, что РФ занимает особое место среди других нервных аппаратов, в значительной мере определяя общий уровень их активности. В первые годы после этих открытий было широко распространено представление, что отдельные нейроны РФ тесно связаны друг с другом и образуют однородную структуру, в которой возбуждение распространяется диффузно. Однако позднее выяснилось, что даже близко расположенные клетки РФ могут обладать совершенно различными функциональными характеристиками. РФ расположена на всем протяжении ствола — от промежуточного мозга до верхних шейных спинальных сегментов. Она представляет собой сложное скопление нервных клеток, характеризующихся обширно разветвленным дендритным деревом и длинными аксонами, часть которых имеет нисходящее направление и образует ретикулоспинальные пути, а часть — восходящие. РФ взаимодействует с большим количеством волокон, поступающим в нее из других мозговых структур — коллатералями проходящих через ствол мозга сенсорных восходящих систем и нисходящими путями, идущими из передних отделов мозга (в том числе из двигательных зон). И те и другие вступают с РФ в синаптические связи. Кроме того, многочисленные волокна поступают к нейронам РФ из мозжечка. Нисходящая часть РФ оказывает неоднозначное влияние на деятельность спинного мозга: раздражение продолговатого мозга (его гигантоклеточного ядра) и некоторых участков варолиева моста сопровождается торможением рефлекторной деятельности нижележащих отделов, а при раздражении более дорсальных и оральных отделов продолговатого мозга — диффузно облегчает действие тех же функциональных структур. Примером первого варианта влияния является эффект мышечного расслабления во время сна. Кроме того, ретикулярное торможение работы нейронов спинного мозга приводит и к ослаблению афферентных восходящих импульсов, то есть снижает передачу сенсорной информации в корковые мозговые центры. Ретикулярные структуры, регулирующие соматические и вегетативные функции, отличаются высокой химической чувствительностью и обнаруживают обратную регулирующую зависимость от характеристик внутренней среды организма (эндокринной системы, уровня СО2 в крови и т. п.). Восходящая часть РФ обеспечивает регуляцию активности высших отделов мозга, главным образом коры больших полушарий. Впервые возможность такого влияния была зарегистрирована в 1935 г. бельгийским нейрофизиологом Бремером в результате перерезки у животных головного мозга на разных уровнях. Поддержание бодрствующего состояния переднего мозга обусловливается первоначальной активацией афферентными раздражениями ретикулярных структур мозгового ствола, а они по восходящим путям определяют функциональное состояние коры, что, конечно, не исключает и прямой передачи афферентации в соответствующие мозговые зоны. Восходящая часть РФ, так же как и нисходящая, помимо деятельности активирующих участков, порождает и общее тормозное влияние. Последнее обеспечивают стволовые участки мозга, в которых найдены так называемые «центры сна», в то время как более дифференцированные по вектору приложения функции предположительно реализуются более высоко расположенными структурами. При исследовании морфологических особенностей клеток РФ было обнаружено, что многие из них имеют Т-образное деление аксонов, один из отростков которых идет вверх, а другой вниз. Это позволило предположить, что и восходящие, и нисходящие функции могут быть связаны с деятельностью одних и тех же нейронов. Кроме того, особенностью отношений коры головного мозга и нижележащих отделов является то, что структуры, обеспечивающие и регулирующие тонус коры, сами находятся с ней в двойных встречных отношениях. Преимущественно тонизируя кору через восходящие пути, лимбические, мезенцефальные и стволовые структуры РФ в то же время подвержены корковой регуляции — и тормозной, и возбуждающей. Эти встречные модулирующие воздействия в первую очередь имеют отношение к лобным отделам, в которых формируются намерения, планы и перспективные программы сознательного поведения. Поведение взрослого человека является примером баланса этих встречных воздействий. РФ не является спонтанно активирующей системой, а берет энергетический потенциал из двух источников — из обменных процессов организма, лежащих в основе гомеостаза, и из поступающих в организм раздражений внешнего мира. Дефицит во внутренней среде стимулирует инстинктивный компонент поведения, а роль второго источника активности может быть проиллюстрирована эффектом засыпания при искусственном отключении основных рецепторных аппаратов (зрения, слуха и кожной чувствительности). С точки зрения психических функций энергетический неспецифический блок имеет отношение к процессам общего и селективного внимания, а также к сознанию в целом, процессам неспецифической памяти (запечатлению, хранению и переработке разномодальной информации), к сравнительно элементарным эмоциональным состояниям (страха, боли, удовольствия, гнева). В исполнении последней функции особую роль играют лимбические отделы мозга, которые помимо эмоционального фона обеспечивают переработку интероцептивной информации. Многими учеными РФ рассматривается как водитель многих биологических ритмов организма, часть из которых не только навязывается извне, но и может поддерживаться без видимой внешней стимуляции. Специфическую роль в этой мозговой системе играет не только афферентный энергетический потенциал, но и информационный аспект раздражителя, выражающийся в категориях сенсорного потока и рационального значения (ценности) стимула. В концентрированной форме специфика работы 1-го блока прослеживается в организации ориентировочного рефлекса: энергетическая мобилизация организма порождается появлением нового стимула, требующего к себе экстренного внимания и сличения с имеющимися в памяти старыми раздражителями, а также последующим переводом полученных итогов в плоскость эмоциональных категорий «вредности-полезности». 2-й блок — приема, переработки и хранения экстероцептивной информации — включает в себя центральные части основных анализаторных систем: зрительной, слуховой и кожно-кинестетической, корковые зоны которых расположены в затылочных, теменных и височных долях мозга (рис. 17). В системы этого блока формально включаются и центральные аппараты вкусовой и обонятельной рецепции, но у человека они настолько оттеснены представительствами высших экстероцептивных анализаторов, что занимают в коре головного мозга незначительное место.
Рис. 17. Функциональные блоки мозга — 2-й блок (по Лурия). Основу данного блока составляют первичные или проекционные зоны коры (поля), выполняющие узкоспециализированную функцию отражения только стимулов одной модальности. Их задача — идентифицировать стимул по его качеству и сигнальному значению, в отличие от периферического рецептора, который дифференцирует стимул лишь по его физическим или химическим характеристикам. Основная функция первичных полей — тончайшее отражение свойств внешней и внутренней среды на уровне ощущения. Все первичные корковые поля, как это было показано с помощью электростимуляции еще канадским нейрохирургом Пенфильдом, характеризуются топическим принципом организации, согласно которому каждому участку рецепторной поверхности соответствует определенный участок в первичной коре («точка в точку»), что и дало основание назвать первичную кору проекционной. Величина зоны представительства того или иного рецепторного участка в первичной коре зависит от функциональной значимости этого участка, а не от его фактического размера. К числу первичных относятся 17-е (для зрения), 3-е (для кожно-кинестетической чувствительности) и 41-е (для слуха) поля. Вторичные поля представляют собой клеточные структуры, морфологически и функционально как бы надстроенные над проекционными. В них происходит последовательное усложнение процесса переработки информации, чему способствует предварительное прохождение афферентных импульсов через ассоциативные ядра таламуса. Вторичные поля обеспечивают превращение соматотопических импульсов в такую функциональную организацию, которая на уровне психики эквивалентна процессу восприятия. На поверхности мозга вторичные поля граничат с проекционными или окружают их. Номера вторичных полей — 18-е, 19-е, 1-е, 2-е, 42-е, 22-е и частично 5-е. Первичные и вторичные поля относятся к ядерным зонам анализаторов. Третичные (ассоциативные) поля (зона перекрытия) имеют наиболее сложную функциональную нагрузку. Они расположены на границе затылочного, височного и заднецентрального отделов коры и не имеют непосредственного выхода на периферию. Их функции почти полностью сводятся к интеграции возбуждений, приходящих от вторичных полей всего комплекса анализаторов. Работа этих зон своим психологическим эквивалентом имеет сценоподобное восприятие мира во всей полноте и комбинации пространственных, временных и количественных характеристик внешней среды, но не исчерпывается этим. Второе значение зон перекрытия — это переход от непосредственного наглядного синтеза к уровню символических процессов, благодаря которым становится возможным осуществление речевой и интеллектуальной деятельности. Третичные поля находятся вне ядерных зон. Особого выделения требует зона ТРО (от латинских названий долей: височной — temporalis, теменной — parietalis, затылочной — occipitalis), которая реализует наиболее сложные интегративные функции — 37-е и частично 39-е поле. Работа второго блока подчиняется трем законам. Закон иерархического строения. Первичные зоны являются фило- и онтогенетически более ранними. Поэтому недоразвитие первичных полей у ребенка приводит к потере более поздних функций (принцип «снизу-вверх»), а у взрослого с полностью сложившимся психологическим строем третичные зоны управляют работой подчиненных им вторичных и при повреждении последних оказывают на их работу компенсирующее влияние (принцип «сверху-вниз»). Выготский следующим образом характеризует данное теоретическое положение: «Объяснение этой закономерности лежит в том факте, что сложные отношения между различными церебральными системами возникают как продукт развития и что, следовательно, в развитии мозга и в функционировании зрелого мозга должна наблюдаться различная взаимная зависимость центров: низшие центры, служащие в истории развития мозга предпосылками для развития функций высших центров, являющихся вследствие этого зависимыми в своем развитии от низших центров, в силу закона перехода функций вверх сами оказываются в развитом и зрелом мозгу несамостоятельными, подчиненными инстанциями, зависящими в своей деятельности от высших центров. Развитие идет снизу вверх, а распад — сверху вниз». Закон убывающей специфичности. Наиболее модально специфичными (в данном случае — ориентированными на конкретное свойство объекта, улавливаемое конкретным видом анализатора) являются первичные зоны. Третичные зоны вообще надмодальны. Закон прогрессирующей латерализации. По мере восхождения от первичных к третичным зонам возрастает дифференцированность функций левого и правого полушария (в основном — по центральным предпосылкам речи и доминантности одной из рук). 3-й блок — программирования, регуляции и контроля за протеканием психической (сознательной) деятельности включает моторные, премоторные и префронтальные отделы коры лобных долей мозга — кпереди от передней центральной извилины (медиобазальные отделы лобных долей входят в первый блок) (рис. 18). Основная цель работы этого блока — формирование планов действий, то есть создание программы психического акта и развертка последовательности исполнения его во времени в реальном поведении. Находясь под постоянным влиянием второго блока, префронтальные (по характеру и сложности обработки информации — ассоциативные) отделы лобных долей одновременно зависимы от речевого и мотивационного компонентов. Подготовка двигательных импульсов завершается их выходом на периферию через двигательную зону коры — преимущественно 4-е поле.
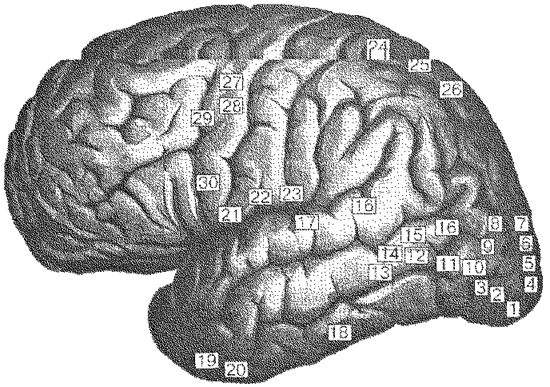
Рис. 18. Функциональные блоки мозга — 3-й блок (по Лурия). Иллюстрацией специфики работы первичных, вторичных и третичных полей указанных блоков является схема галлюцинаций, вызываемых искусственными раздражениями различных участков мозга (рис. 19).
Рис. 19. Схема искусственно вызываемых эффектов при раздражении коры головного мозга (по Лурия): 1— светящиеся шары; 2 — окрашенный свет; 3 — белый свет; 4 — голубые диски с красными кольцами; 5 — пламя, надвигающееся со стороны; 6 — голубой туман, надвигающийся со стороны; 7 — лица, звери, идущие со стороны; 8 — друг, идущий со стороны и делающий знак; 9 — лица; 10 — звери; 11 — лица и бабочки; 12 — полет птиц; 13 — желтый цвет; 14 — лица и животные вниз головой; 15 — человеческие фигуры; 16 — фигуры; 17 — шум, голоса; 18 — шум; 19, 20 — шум барабана; 21, 22 — вкусовые галлюцинации; 23 — ощущение движения в языке; 24, 25, 26 — вестибулярные галлюцинации; 27, 28, 29 — насильственно издаваемые звуки; 30 — насильственное издавание слов. Понятие нейропсихологического фактора и синдрома. Фактор (вообще) — движущая сила совершающегося процесса или одно из его необходимых условий. Нейропсихологический фактор — принцип физиологической деятельности определенной мозговой структуры. Он является связующим понятием между психическими функциями и работающим мозгом. С одной стороны, фактор является результатом активности определенных функциональных органов мозга, а с другой — играет объединительную роль для психических процессов в их системной функции реализации какого-либо специфического звена. Поражение той или иной мозговой структуры (одного из компонентов функциональной системы) может проявляться в полном или частичном выпадении ее функции, либо в патологическом изменении режима ее деятельности (угнетении, раздражении, смены принципа работы). То общее, что может быть обнаружено в изменениях, регистрируемых при выпадении или искажении каких-либо физиологических или «обслуживаемых» ими психических функций, по сути и есть изменения нейропсихологического фактора. Фактор — это обобщенный и связанный с определенным динамически локализованным нейронным ансамблем смысл его работы. Инструментом выделения нейропсихологических факторов является синдромный анализ, включающий три этапа осмысления регистрируемых изменений психической деятельности. 1. Качественная квалификация нарушений психических функций с объяснением причин возникших изменений. 2. Анализ и сопоставление первичных и вторичных расстройств, то есть установление причинно-следственных связей между непосредственным источником патологии и возникающими по закону системной организации функций производными расстройствами. В частных случаях таким последствием может быть полный распад соответствующей психической функции. К числу третичных нарушений иногда относят компенсаторные перестройки той или иной функциональной системы в ответ на возникшее поражение с целью замещения пораженного звена. 3. Изучение состава сохранных ВПФ, облегчающее дифференциальную топическую диагностику. В отношении детского возраста психологическая оценка нарушений развития или локальных поражений мозга не может быть полной, если она не учитывает также и отклонений от среднего возрастного развития, на котором находится ребенок, или особенностей дизонтогенеза (расстройства индивидуального развития), вызванного болезненным процессом или его последствиями. Различные виды психического дизонтогенеза ребенка могут обусловливаться как автономным влиянием биологических или социальных факторов, так и их сложной комбинацией, отличающейся по механизмам воздействия на различных этапах развития организма. Выготский выделял следующие факторы, определяющие аномальное развитие. Время возникновения первичного дефекта. Общим для всех видов аномального развития является раннее возникновение первичной патологии. Дефект, возникший в раннем детстве, когда не сформировалась вся система функций, обусловливает наибольшую тяжесть вторичных отклонений. Из-за системного строения психики вторичные отклонения становятся причиной недоразвития других психических функций. Например, недоразвитие слуха может приводить к нарушениям речевых функций, а те, в свою очередь — к нарушению интерперсональных отношений. Чаще повреждаются подкорковые функции, имеющие короткий цикл развития в онтогенезе. Корковые функции, имеющие более длительный период развития, при раннем вредном воздействии чаще страдают или временно задерживаются в своем развитии. Степень выраженности первичного дефекта. Различают два основных вида дефекта. Первый из них — частный, обусловленный дефицитом отдельных функций гнозиса, праксиса, речи. Второй — общий, связанный с нарушением регуляторных систем. Чем глубже первичный дефект, тем больше страдают другие функции. Указывают на два основных типа дизонтогенеза — ретардацию и асинхронию (Ушакова и Ковалева). Под ретардацией понимают запаздывание или остановку психического развития. Нейрофизиологической основой парциальной (частичной) ретардации является нарушение темпов и сроков созревания отдельных функциональных систем. Характерным признаком асинхронии является выраженное опережение в развитии одних психических функций и свойств формирующейся личности и значительное отставание темпов и сроков созревания других функций и свойств. Это становится основой дисгармоничного развития психики в целом. Следует отличать асинхронию от физиологической гетерохронии, то есть разновидности созревания церебральных структур и функций, что наблюдается при нормальном психическом развитии. Особо рассматривают третий тип дизонтогенеза, в основе которого лежит преходящая физиологическая незрелость, а также временный возврат к незрелым формам нервно-психического реагирования у ребенка. Чрезвычайно важен анализ связи фиксируемых симптомов с критическими или сенситивными периодами в развитии функциональных систем, гетерохронностью созревания мозга, особенностью вертикальных, внутриполушарных и межполушарных взаимодействий, промежуточными и окончательными стадиями формирования наиболее поздно созревающих ассоциативных структур мозга. Для детского мозга в целом характерна более высокая пластичность, вследствие чего нейропсихологические симптомы отчетливо проявляются лишь при бурно развивающихся патологических процессах или непосредственно в ближайшие сроки после мозговых поражений. Из-за несовпадения концепций различных авторов и сложности самого понятия нейропсихологического фактора, при попытках классифицировать многообразный исходный эмпирический материал возникают различные варианты основных смысловых нагрузок, составляющих содержание конкретного фактора. В качестве относительно самостоятельных могут быть рассмотрены следующие. Модально-неспецифический (энергетический) фактор — связан с работой глубинных отделов мозга. Они закладываются еще во время внутриутробного развития, но обеспечить стабильность баланса возбуждения и торможения в первые годы жизни еще не могут. Дозревание этих церебральных отделов продолжается после рождения. Феноменологически неспецифический фактор выражается степенью активности мозговых структур в континууме «сон-бодрствование», что предполагает участие прежде всего отделов, имеющих отношение к витальным потребностям и жизнеобеспечению организма (на уровне темперамента в данном контексте принято говорить о стеничных и астеничных людях). В этих же рамках реализуется управление биологическими ритмами. Черепно-мозговые травмы, даже протекающие без видимых симптомов, недостаточность в работе сердечно-сосудистой системы, неблагоприятные экологические и метеорологические влияния, интоксикации различного рода, последствия инфекционных заболеваний относятся к биологическим причинам, снижающим уровень активности глубинных структур и мозга в целом. К аналогичным по результатам психогенным предпосылкам относятся стрессовые ситуации, длительные переживания (особенно хронические конфликты), недозированные психические нагрузки. Эти обстоятельства приводят к нарушению гармонии в смене фаз активности и бодрствования, повышенной утомляемости, замедленному включению в деятельность, колебаниям ее продуктивности, расстройствам сна. Наиболее отчетливо слабость фактора энергетического обеспечения обнаруживается в процессах памяти и внимания: запомненное быстро забывается, особенно после выполнения какого-либо действия после запоминания (следы «затираются» по принципу интерференции); внимание становится колеблющимся, встречаются трудности его распределения и переключения, возникает повышенная истощаемость к концу выполнения задания. Кинетический фактор — связан с работой ассоциативных премоторных отделов мозга и обеспечением такой составляющей психической деятельности, как возможность естественного и целесообразного перехода от одного элемента к другому при выполнении различных действий — цепи последовательно сменяющих друг друга шагов. В реальной жизни к таким действиям относятся разнообразные двигательные акты, осуществляемые в форме кинетических (мелодических) схем. При нарушениях данного фактора в случае мозговой патологии или при его несформированности у детей, элементы движений выполняются изолированно, двигательный цикл характеризуется прерывистостью, затрудняется быстрая и плавная смена включенных в движение компонентов. Особенно ярко эти черты обнаруживаются в моторном обеспечении письма и рисования. В более выраженных случаях недостаточности кинетического фактора могут возникать своеобразные застревания на каком-то фрагменте движения, приводящие к его неоднократным повторениям. В письме это проявляется в неконтролируемых повторах букв и их частей, особенно в тех случаях, когда буквы содержат сходные по написанию элементы. В графических действиях каждая линия вырисовывается отдельно или воспроизводится многократно в виде штрихов. Становится невозможным остановить ранее начатое движение. Все отмеченные аномальные механизмы относятся и к речевой моторике, поскольку она требует плавной смены артикуляции и перехода от слова к слову при построении высказывания: пропускаются согласные в тех словах, где они сочетаются («страшный-срашный»), а также появляется телеграфный стиль с преимущественным употреблением существительных в именительном падеже или глаголов в неопределенной форме. Предполагается, что такие особенности речи могут быть связаны не только с ее внешней, собственно моторной составляющей, но и с последовательным развертыванием смысловой схемы высказывания, представленной во внутреннем плане. Динамика мыслительного процесса также теряет свою плавность. Это может проявляться в замедленном понимании арифметических задач, в необходимости многократного прочитывания условий, в счетных операциях, которые плохо автоматизируются. Слабость кинетического фактора обнаруживается и при решении логических задач, в которых необходим переход к новому алгоритму решения. Популярное:
|
Последнее изменение этой страницы: 2016-03-22; Просмотров: 1198; Нарушение авторского права страницы