
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Объем и быстродействие памяти
Если следовать логике Д. Гартли, А. А. Ухтомского, А. Ф. Самойлова, М. Н. Ливанова, Г. Уолтера, Е. Р. Джона, К. Прибрама и других сторонников идеи динамического кодирования воспринимаемой и хранимой в памяти информации, то можно предположить, что нейронные ансамбли, ответственные за субъективное отражение, активируются периодически, разряжаясь импульсами. Из-за биений частот, слагающих ЭЭГ, актуализированные образы памяти как бы пульсируют с периодом биений, максимальная длительность которого вычисляется по следующей формуле: Т = 1/(FR). Заметим, что 1/R = FT. Из всего набора С образов долговременной памяти в каждый текущий момент актуализировано ограниченное число М разных образов [Atkinson et al, 1974]. В каждый текущий момент с вероятностью 1/М один из образов имеет максимальную возбудимость. Время реакции в ответ на появление стимула, адекватного образу, в этот момент минимально. Стимулы подаются независимо от периодических колебаний активности нейронов (экспериментатор их не видит). Следовательно, вероятность совпадения стимула с той или иной фазой колебаний возбудимости одинакова в течение всего периода колебаний. Какие-то стимулы совпадают с фазой повышенной возбудимости, и ответные реакции следуют без всякой дополнительной задержки. В остальных случаях задержка равномерно распределяется в течение всего отрезка пониженной возбудимости ансамблей нейронов. Сказанного достаточно для расчета средней длительности задержки t в зависимости от числа М равновероятно ожидаемых и числа К одновременно предъявляемых стимулов:
где р = (1 - (1 - R)/KM); T= 1/FR; F = 10 Гц (константа Бергера); R = 0, 1 (константа Ливанова). Уравнение выражает количественно скорость обработки информации человеком. В частности, время, необходимое в среднем для опознания одного стимула из числа М равновероятных стимулов, определяется по следующей формуле:
В психологии существует закон скорости обработки информации человеком, установленный У. Хиком [Hick, 1952]. Время обработки прирастает линейно при линейном росте логарифма от количества альтернатив в ситуациях выбора. Основной недостаток этого закона — узкая сфера его действия. Закон справедлив, если количество альтернатив меньше десяти. Закон подвергался критике и был предметом многих дискуссий. Уравнение (1), включающее в себя обе физиологические константы и выведенное из представлений о кодировании информации циклами нейронной активности, намного точнее. Оно действенно при неограниченном количестве альтернатив и предсказывает результаты психологических опытов с высокой степенью точности [Бовин, 1985]. В исследованиях И. Ю. Мышкина, А. В. Пасынковой, Ю. А. Шпатенко, Т. С. Князевой, Г. В. Котковой, Д. В. Лозового, О. Ж. Кондратьевой, В. К. Оше и других сотрудников лаборатории А. Н. Лебедева было установлено, что уравнение (1) для оценки быстродействия восприятия и памяти правильно предсказывает разнообразные психологические данные. Следовательно, представление о кодировании воспринимаемых и хранимых в памяти сведений волнами нейронной активности имеет солидное экспериментальное основание.
С одной стороны, память человека выглядит безбрежной. Это долговременная память. С другой — удивительно маленькой. Такова оперативная, или кратковременная (рабочая, как ее иногда называют), память. А раньше ее называли объемом сознания. Психологи попробовали решить задачу о зависимости объема кратковременной памяти от алфавита запоминаемых стимулов и отступились. Появилось правило Д. Миллера «семь плюс или минус два», утверждающее независимость объема от алфавита запоминаемых стимулов [Miller, 1956]. Заданный разброс широк, но на деле он еще больше: от одной-двух единиц (например, в случае иероглифов) до 25-30 в случае бинарных сигналов. Представление о циклах нейронной активности как материальном субстрате памяти и здесь оправдало себя в полном соответствии с исходной идеей Д. Гартли. Единицами памяти, ее нейронными кодами служат пакеты волн, т. е. синхронных импульсных разрядов многих нейронов в составе одного ансамбля. Нейронных ансамблей огромное множество. Каждый из них хранит информацию о каком-то объекте памяти в виде устойчивого волнового узора. Ансамбль состоит из нескольких групп нейронов. Отдельная группа способна генерировать последовательно от 1 до 10 когерентных залпов импульсов за один период доминирующих колебаний при условии, что интервалы между залпами не меньше, чем ливановская ступенька R = 0, 1 по отношению к длительности доминирующего периода. Число нейронов в ансамбле варьирует. Чем больше нейронов вовлекается в ритмы какого-то ансамбля, тем выше вероятность осознания соответствующего образа. Минимальное количество нейронов, обеспечивающее устойчивость ансамбля, порядка 100-300, согласно расчету А. Н. Лебедева и В. А. Луцкого (цит. по [Забродин, Лебедев, 1977, с. 243]. Не синапсы и даже не отдельные нейроны типа нейронов-детекторов или командных нейронов служат единицами хранения, а лишь группы, ансамбли содружественно пульсирующих нейронов. Разумеется, это не атомные и не молекулярные, а клеточные, именно нейронные коды. Их можно назвать также циклическими кодами памяти, потому что цикличность, т. е. регулярность разрядов массы нейронов, отражаемая в регулярности волн электроэнцефалограммы, является специфической особенностью таких кодов. Алфавит нейронных единиц памяти легко рассчитывается. Он находится в обратной зависимости к константе Ливанова. А именно, один из множества залпов указывает на начало периода. Именно поэтому размер алфавита таких кодовых единиц на единицу меньше, т. е. N = 1/R - 1. Число нейронных групп, вовлекаемых в активное состояние за один период (последовательно друг за другом) равно тому же числу, N = 1/R - 1. Как мы видим, длина кодовых цепочек, т. е. последовательно вовлекаемых нейронных ансамблей, лимитирована той же самой частотной рефрактерностью и так же легко рассчитывается. Отсюда выводится максимально возможное количество разных кодовых последовательностей (около полумиллиарда) по формуле: С = NN = (1/R – l)(1/R-1). Такова емкость долговременной памяти, С = 99 = 387 420 489 единиц памяти. Каждая единица памяти — это одно определенное понятие или команда, т. е. паттерн действия. Приведем сравнение: размер активного словаря на родном языке около 10 000, и даже у Шекспира и Пушкина, словарь произведений которых подсчитан, он меньше 100 000 слов. Следовательно, человек способен владеть десятками языков, что, конечно, не ново. Ново то, что емкость памяти — функция одной-единственной физиологической константы (R = 0, 1). Это дробь Ливанова (она названа так по аналогии с другой знаменитой константой — дробью Вебера) (см. след, параграф). Вычисленная емкость позволяет узнать зависимость объема кратковременной памяти от алфавита запоминаемых стимулов. В одном уравнении мы связали три фундаментальных психологических показателя: емкость долговременной памяти (С), емкость оперативной, или рабочей памяти (Н), и емкость внимания (М), т. е. число актуализированных различных образов долговременной памяти:
где А £ М £ А´ Н, и, в свою очередь, С = (1/R - 1)(1/R-1), где R — физиологическая константа Ливанова (R = 0, 1); А - размер заданного алфавита стимулов. Следует пояснить еще раз, что не все единицы памяти, т. е. не все ансамбли актуализированы одновременно. Только небольшое число М ансамблей актуализировано в каждый текущий момент времени. Это число служит мерой объема внимания. Если человек сосредоточил внимание в определенный момент времени на запоминании двоичных элементов (нулей и единиц), то наименьший объем внимания равен размеру объективно заданного двоичного алфавита, знакомого ему, т. е. М = А = 2, Наибольший объем внимания равен следующему произведению: М = А ´ H (в данном примере М = 2 ´ H, где H — коэффициент пропорциональности, равный объему кратковременной, или рабочей, памяти для запоминаемых элементов; это максимальное значение коэффициента). Кратковременная память H измеряется максимальным числом элементов, не обязательно разных и правильно воспроизведенных с учетом их значения и позиции в ряду после однократного восприятия. Длительность однократного восприятия не превышает 2-10 с. Из уравнения (2) следует простое правило для прогноза емкости кратковременной памяти на комбинации признаков, если измерены емкости на каждый из признаков в отдельности:
где Н— искомый объем для комбинации; Н1, Н2, Н3 — объемы кратковременной памяти на исходные признаки. Эта формула, выведенная аналитически из предыдущей, предсказала существование нового феномена, ранее неизвестного в психологии (кроме того, с высокой степенью точности). Ошибка прогноза в разных опытах Н. А. Скопинцевой, Л. П. Бычковой, М. Н. Сыренова и других исследователей по проверке формулы (3) нередко составляла всего 3-5 %. Сравните этот показатель с 25-35 % по правилу Миллера, которое в этой ситуации работает неудовлетворительно. По Миллеру, такая задача неразрешима. В работах И. Ю. Мышкина и В. В. Майорова [Мышкин, Майоров, 1993], плодотворно развивающих теорию динамической памяти, а также в других исследованиях [Маркина и др., 1995] были установлены искомые зависимости объема памяти от параметров электроэнцефалограммы. Таким образом, была реализована цель И. П. Павлова — количественно описать известные психологические явления и предсказать новые с помощью физиологических понятий (причем фундаментальные психологические явления, описывающие объем памяти и ее быстродействие). Примечательно, что в уравнения для расчета емкости памяти человека и ее быстродействия входят два параметра ЭЭГ, частотная рефрактерность (R) и доминирующая частота (F). Они являются, как принято говорить после П. К. Анохина, системообразующими параметрами, должными объяснить множество психологических показателей. Уравнения (1), (2) вместе с их выводом и экспериментальной проверкой детально рассмотрены в некоторых работах [Лебедев, 1982; Лебедев и др., 1985]. Найденные физиологические формулы памяти и ее быстродействия обеспечили решение двух старинных психологических проблем. Нас интересует, в первую очередь, проблема моментального выбора, поиска нужных сведений в памяти, сведений, необходимых на каждом шаге для реализации целенаправленного поведения. В когнитивной психологии, пожалуй, больше всего литературы по парадигме С. Стернберга, ученика Д. Луса, касающейся скорости поиска сведений в памяти [Sternberg, 1969]. С. Стернберг придумал методику для определения такой скорости. Выявилась яркая зависимость скорости от размера ряда запомненных стимулов. П. Каванох [Cavanagh, 1972] обработал данные множества исследователей и обнаружил константу около. 1/4 с, характеризующую время сканирования всего содержимого кратковременной памяти независимо от содержания запомненного материала. По методике С. Стернберга, человек сначала запоминает ряд стимулов, например цифр, как целое, — как единичный образ, — и удерживает этот новый образ до момента появления одного-единственного стимула, который входит в запомненный набор (или, напротив, не входит в него), отвечая нажатием на соответствующую клавишу. В этом случае, по условиям опыта, параметр М из уравнения (1) равен объему Н кратковременной памяти, а параметр К = 1.
Вычисленная из физиологических данных величина отличается от опытного значения, определенного Каванохом по множеству психологических данных, менее чем на 3 %. Интересно заметить, что при Н = 1 (разумеется, по условиям измерения К = 1) время сличения по формуле (1) минимально (около 5 мс). Оно равно константе Гайсслера, с точностью до 0, 3 мс [Geissler, 1990]. Для оценки среднего прироста времени при H > 1 в расчете на один стимул следует разделить найденное значение 0, 5(H + 1)t/H времени сканирования всего содержимого кратковременной памяти на количество приращений (H-1) стимульного ряда. Психологические данные полностью согласуются с физиологическим расчетом [Лебедев и др., 1985; Lebedev, 1990]. Еще одно предсказание касается скорости зрительного поиска, также следующей чисто аналитически из уравнения (1). Формула (1) не только устанавливает зависимость скорости поиска от индивидуальных электрофизиологических констант, но и от размера алфавита воспринимаемых зрительных сигналов [Лебедев и др., 1985]. Из-за циклических колебаний возбудимости нейронных ансамблей образы долговременной памяти, в том числе образы воспринимаемых и произносимых слов, актуализируются не все сразу, а по очереди, причем некоторые чаще, другие реже. По частоте актуализации, т. е., например, по частоте появления одного и того же слова в письменной речи, можно судить о закономерностях циклических нейронных процессов и, наоборот, по особенностям нейронных циклов предсказывать характеристики речи. Если моменты актуализации разных образов совпадают, то такие единицы памяти имеют шанс объединиться. Таким образом вырабатывается новое понятие. Так происходит научение и реализуются акты творчества. Выживают, т. е. не объединяются навсегда в одном ансамбле, только те образы памяти, циклическая активность которых не коррелирует между собой. Периоды циклов такой активности соотносятся как члены натурального ряда 1: 2: 3: 4..., а вероятности актуализации, как члены гармонического ряда (1/1): (1/2): (1/3): (1/4). Сумма вероятностей равна единице, а значение первого члена равно физиологической константе Ливанова. Так выводится следующая формула, с помощью которой можно предсказать зависимость частоты появления слова (р) в связной речи от номера его ранга: р - R/i, (4) где i — ранг слова по частоте появления в тексте. Формула, включающая в себя физиологическую константу, выражает собой известный с 1930-х гг. закон Ципфа [Zipf, 1935]. Из формулы (4) следуют уравнения для расчета зависимости объема словаря от размера текста, в котором данный словарь реализован, и вычисления интервалов между повторениями одного и того же слова в тексте [Лебедев, 1985]. Речь, письменная или устная, причем не только поэзия, музыкальна. Константа Ливанова входит в уравнение (4) гармонического ряда слов, ранжированных по частоте. Используя уравнения множественной линейной регрессии для оценки способности школьников к обучению по характеристикам ЭЭГ, мы нашли, что параметры альфа-ритма, определяющие емкость памяти, влияют также на успешность прогноза интеллектуального развития [Артеменко и др., 1995], что неудивительно. Таким образом, теория циклических нейронных кодов памяти позволяет по-новому взглянуть на уже известные психологические законы.
Диапазон ощущений
В психофизике дробь Вебера определяет границу между ощущаемым и неощущаемым. Это едва заметный субъективно прирост ощущения, взятый по отношению к исходной величине стимула. Вытекающая из дроби оценка величины ощущений — одна из центральных проблем психологии. Недаром закон, открытый Г. Фехнером, во всех энциклопедиях называют основным законом психофизики. До сих пор не затухают жаркие дискуссии по поводу основного психофизического закона в двух его ипостасях: формулировке его первооткрывателя Г. Фехнера о логарифмической зависимости силы ощущения от интенсивности стимула и утверждении С. Стивенса о степенном характере такой зависимости [Забродин, Лебедев, 1977]. Опираясь на данные французского психолога А. Пьерона о нелинейной зависимости времени простой реакции от интенсивности стимула, мы находим физиологическое решение проблемы основного психофизического закона. А. Пьерон обнаружил, что скрытое время простой сенсомоторной реакции обратно связано с интенсивностью стимула, возведенной в некую степень. Позже было обнаружено, что показатель степени имеет ту же величину, что и показатель степени в основном психофизическом законе Стивенса [Bonnet, 1990] для яркости и громкости. Следовательно, произведение скрытого времени t на величину ощущения S, измеренного по Стивенсу, есть константа, выраженная в условных единицах: tS = 1 (5) Диапазон всевозможных ощущений задан константой С = NN согласно формуле (1). Минимальная перцептивная задержка вычисляется по формуле (1) при К = 1 и М = 1. Она равна 5 мс, что соответствует константе Гайсслера (Geissler, 1990). Максимальная задержка для слабых, околопороговых сигналов при К > 1 и М > > 1 приближается к 1000 мс. Отношение максимальной задержки к минимальной (1000/5 = 200) соответствует диапазону ощущений. Минимальное значение показателя степени (Kmit) в законе Стивенса находится по формуле Kmin = log 200/log (С), (6) где С — объем долговременной памяти, вычисленный ранее. Остальные значения показателя (Ki) вычисляются по той же формуле с подстановкой в нее логарифма отношения верхнего (болевого) порога интенсивности для i-й модальности (зрительной и слуховой) к нижнему абсолютному порогу вместо выражения log(C). Опытные данные многих авторов, обобщеннные Р. Тетсуняном [Teghtsoonian, 1971], подтверждают точность подобного расчета. Отношение (t1 — t2)/t1 выражает характерную особенность дроби Вебера, в которой t1 и t2 — скрытые задержки восприятия, вычисленные по формуле (5) при подстановке в нее ощущений (S) в относительных единицах (от 1 до 200), вызванных едва различимыми по интенсивности стимулами. Последовательно уменьшающиеся значения дроби равны членам гармонического ряда 1; 0, 5; 0, 33; 0, 25 и т. д. Дробь Вебера, надо заметить, также служит предметом незатухающих дискуссий [Lebedev, 1993b; Link, 1992]. Оригинальную ее интерпретацию предложил недавно Е. Н. Соколов в докладе, посвященном памяти Э. Г. Вебера. Таким образом, из физиологических предпосылок вытекает органическая связь временных параметров восприятия стимулов с ощущением их интенсивности. Показатель степени в законе Стивенса (6) является сложной функцией нейрофизиологических параметров. Чем больше разнообразие нейронных кодов, тем шире диапазон ощущений.
Нейронные коды памяти
В уравнениях (1)—(6) с электрофизиологическими константами нашла выражение упомянутая ранее мечта И. П. Павлова — количественно объяснить известные психологические явления и предсказать новые с помощью физиологических показателей. Впервые по физиологическим показателям благодаря трудам Г. Бергера и М. Н. Ливанова вычислены объем памяти и ее быстродействие. Весьма вероятно, что на этой основе будут созданы новые эффективные методы обучения с использованием компьютеров, воспринимающих потенциалы мозга, и разработаны новые поколения нейрокомпьютеров, способных к творчеству и далекому прогнозу событий. Компьютерные сети, которые распространяются по всему миру, уподобятся человеческому мозгу и превзойдут человеческий интеллект. Несомненно, наступит эпоха самоорганизации и «кристаллизации» информации, заключенной в гигантских нейроноподобных сетях. То, что теория условных рефлексов была названа учением, т. е. непогрешимой истиной, нельзя ставить в вину И. П. Павлову. Теория Павлова — одна из первых ступенек на лестнице, ведущей к преобразованию психологии в точную науку по пути, ранее пройденному физиками и химиками. В этом ее смысл. В заключение еще раз подчеркнем, что крупной вехой в движении по заданному Павловым направлению стали работы академика М. Н. Ливанова. Две физиологические константы (Ливанова, R = 0, 1, и Бергера, F = 10 Гц) в уравнениях когнитивной психологии количественно объясняют фундаментальные психологические особенности обработки информации человеком, включая закономерности восприятия, памяти и речи. Константа Ливанова ограничивает разнообразие циклических кодов памяти, а вместе с константой Бергера — также и ее быстродействие.
Глава 7 ПСИХОФИЗИОЛОГИЯ ЭМОЦИЙ
Популярное:
|
Последнее изменение этой страницы: 2016-03-26; Просмотров: 628; Нарушение авторского права страницы
 (1)
(1)
 Теперь перейдем от временных характеристик восприятия к оценке объемов воспринимаемой и хранимой в памяти информации. Психологи давно уже выделили несколько типов памяти человека: иконическую, кратковременную и долговременную. Имеются и другие классификации.
Теперь перейдем от временных характеристик восприятия к оценке объемов воспринимаемой и хранимой в памяти информации. Психологи давно уже выделили несколько типов памяти человека: иконическую, кратковременную и долговременную. Имеются и другие классификации.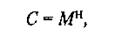 (2)
(2) (3)
(3) Для сличения одного образа стимула с предъявленным требуется t/H времени, а для опознания предъявленного стимула, если его образ присутствует в запомненном ряду, требуется в сумме от 1 до числа Н сличений, в среднем (1 + Н)/2 сравнений, т. е. 0, 5(Н + 1)t/H единиц времени, что равно 0, 25 с при типичных значениях F = 10Гц и R = 0, 1.
Для сличения одного образа стимула с предъявленным требуется t/H времени, а для опознания предъявленного стимула, если его образ присутствует в запомненном ряду, требуется в сумме от 1 до числа Н сличений, в среднем (1 + Н)/2 сравнений, т. е. 0, 5(Н + 1)t/H единиц времени, что равно 0, 25 с при типичных значениях F = 10Гц и R = 0, 1. Нейронными кодами памяти служат циклически повторяющиеся волны импульсов, порождаемые нейронными ансамблями. Длительность одного цикла составляет около 100 мс. Заметим, что по данным А. М. Иваницкого, работавшего с вызванными потенциалами, примерно столько же времени уходит на формирование перцептивного образа [Иваницкий, 1976]. Число ансамблей, последовательно вовлекаемых в активность за этот период, определяется константой Ливанова. Чаще всего оно равно 10. Каждый ансамбль за один цикл активности генерирует от 1 до 10 залпов из нейронных импульсов множества нервных клеток, образующих ансамбль. Минимальное число нейронов, обеспечивающее устойчивые, незатухающие колебания ансамблевой ативности, составляет около 100 клеток. Максимальное число неизвестно (порядка десятков и сотен тысяч). Указанные числовые значения зависят от индивидуальных значений двух электрофизиологических констант — Ливанова (около 1/10 от цикла активности) и Бергера (порядка 10 Гц).
Нейронными кодами памяти служат циклически повторяющиеся волны импульсов, порождаемые нейронными ансамблями. Длительность одного цикла составляет около 100 мс. Заметим, что по данным А. М. Иваницкого, работавшего с вызванными потенциалами, примерно столько же времени уходит на формирование перцептивного образа [Иваницкий, 1976]. Число ансамблей, последовательно вовлекаемых в активность за этот период, определяется константой Ливанова. Чаще всего оно равно 10. Каждый ансамбль за один цикл активности генерирует от 1 до 10 залпов из нейронных импульсов множества нервных клеток, образующих ансамбль. Минимальное число нейронов, обеспечивающее устойчивые, незатухающие колебания ансамблевой ативности, составляет около 100 клеток. Максимальное число неизвестно (порядка десятков и сотен тысяч). Указанные числовые значения зависят от индивидуальных значений двух электрофизиологических констант — Ливанова (около 1/10 от цикла активности) и Бергера (порядка 10 Гц).