|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ФИЛОГЕНЕЗ СКЕЛЕТА ТУЛОВИЩА И ХВОСТА
Ряд этапов обрисованного индивидуального развития в общих чертах очень напоминает филетическое преобразование скелета туловища. Самым древним, а следовательно, и наиболее примитивным осевым скелетом у предков позвоночных служила спинная струна —chorda dorsalis. У хордовых животных (ланцетник) она является единственным, в достаточной степени прочным и упругим осевым шнуром. У некоторых низших позвоночных—круглоротых, осетровых, двудышащих
рыб—хорда в переживаемую эпоху функционирует в продолжение всей жизни в качестве основной скелетной оси наряду с хрящевыми элементами позвонков. Её клетки с прочными стенками своих вакуолей обладают большим внутренним давлением (тургором); большой толщиной и прочностью отличается и наружная оболочка хорды. В эмбриональной жизни хорда свойственна всем позвоночным, в том числе и млекопитающим. Однако при наличии хорды как единственной более прочной осп у хордовых животных имеется ещё мягкий соединительнотканный остов тела. Он связан с осью и отдаёт в туловище и хвосте ряд поперечных прослоек, к которым прикрепляются мускульные сегменты, обслуживающие движение животных в воде. Хрящевая стадия. Замена хорды более прочным скелетом первоначально свелась к упрочению области её оболочки, где соединительная ткань замещалась окружающими хорду хрящевыми пластинчатыми элементами (рис. 27—А, В), которые в дальнейшем постепенно вытеснили хорду, формируя по оси сегментально расположенные зачатки позвонков. В наиболее полно раз питом виде таких хрящевых пластинок имелось четыре пары. Из них верхняя главная п а р а д у г (С, 2), срастаясь, давала главным образом нервную дужку позвонка (11), т. е. участок позвоночного канала для спинного мозга, над которым слившимися дужками формировался остистый отросток (72). Основания, т. е. вентральные участки, этих дужек участвовали в образовании верхней части тела позвонков, т. е. опорных кусков осевого скелета. Нижняя главная пара дуг (7), срастаясь одна с другой, давала дужку, охватывающую хорду с нижней стороны, и входила у ряда позвоночных в состав развивающегося тела позвонков (13). Два других парных вставочных элемента, меньших по размеру, также предназначались для формирования тела позвонка. Таким образом, хрящевой зачаток тела позвонка был сложным.
Рис. 27. Последовательные стадии образования позвонков: А—у миноги, В—у зародыша селяхии, С—последовательные стадии (I, II, III) образования позвонка селяхий. 1—передняя и 2—задняя дорзальные дуги; 3—миосепты; 4—дорзальный и 5—вентральный нервные корешки; 6—хорда; 61—ее оболочка; 7—вентральные дуги; 8—ребро; 9—гемальный отросток; 10—поперечный отросток позвонка; 11—невральная дуга; 12—остистый отросток; 13—тело позвонка
Костная стадия. У многих позвоночных позвонки так и остаются в хрящевой стадии (хрящевые рыбы), но у большинства переходят в костную стадию. У низших позвоночных с костным скелетом (костные ганоиды и костные рыбы) стенка цилиндрического тела позвонка развивается лишь в его средней части, а передний и задний концы остаются сильно вогнутыми (амфицельная форма), в силу чего хорда сохраняется главным образом между соседними позвонками (рис. 28—А, 1). У земноводных, а именно у низших хвостатых амфибий, тела позвонков также двояко вогнуты, со слабо развитыми межпозвоночными хрящами, а у высших хвостатых амфибий межпозвоночные хрящи выражены несколько сильнее и развились, по-видимому, из вставочных хрящевых элементов. Эти хрящи сильно перетягивают к центру хорду, но нередко она ещё сохраняется внутри тел позвонков в виде незначительного остатка. Тела позвонков имеют вогнутость лишь на переднем своём конце (процельная форма) (Л, 2), а у бесхвостых амфибий — на заднем конце (опистоцельная форма) (А, 3). У млекопитающих прирастающие к телам позвонков эпифизы делают передний конец их выпуклым, а задний—плоским (опистоцельные позвонки) и хорда в виде очень незначительного и сильно изменённого остатка (пульпозное ядро хрящевой прослойки) сохраняется между позвонками. Первично ребро было двойным и стало единым цельным вследствие срастания. У некоторых костистых ганоидов—crossopterygii—на каждый сегмент приходится по две пары рёбер—пара глубоких, так называемых висцеральных, и пара поверхностных — париетальных. Париетальные рёбра залегают в боковой продольной соединительнотканной прослойке (правой и левой стороны), отделяющей дорзальную мускулатуру от вентральной, и находятся в пределах сегмента не только поверхностнее, но и каудальнее глубоких рёбер. Последние помещаются в межмускульных прослойках (миосептах) и топографически занимают место не только глубже, но и краниальнее поверхностных рёбер. Появление грудной кости у позвоночных обусловлено развитием у них ногообразных грудных конечностей как опоры о почву или воздух (птицы); это ясно из того, во-первых, что плечевой пояс, если он полно развит, тесно связывается с грудной костью, а во-вторых, что с редукцией грудной конечности у amniota обыкновенно исчезает и грудная кость (у безногих ящериц). Таким образом, она содействует прочности краниальной части грудной клетки, к которой приставлены грудные конечности.
В период эволюции высших позвоночных скелет туловища и хвоста параллельно с мягкими частями тела претерпел ряд существенных видоизменений. На своём протяжении от затылочной кости до конца хвоста он всё более усложнялся тем, что рельефнее распадался на ряд отделов. Каждый отдел одновременно с диференцировкой пищеварительного и дыхательного аппаратов и поясов конечностей приобретал свои характерные признаки. У наиболее примитивных водных позвоночных (рыбы) осевой скелет состоит лишь из трёх участков: головного, туловищного и хвостового.
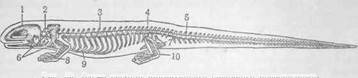
X в о с т о в ы й отдел выделяется благодаря выпавшей на его долю специальной функции передвижения животного в воде. В силу этого отдел хвоста остаётся свободным от внутренностей. У рыб его редуцированные (за отсутствием внутренних органов) висцеральные рёбра смыкаются в каждом сегменте в очень маленькую дугу. Такие дуги охватывают собой лишь проходящий в хвостовом отделе сосуд, вследствие чего они и могут быть названы гемальными дужками. В то же время хвостовый отдел, как главный орган движения вперёд, очень обильно снабжён мускулатурой. Оставшийся собственно туловищный отдел характеризуется присутствием в каждом сегменте, кроме позвонка, одной (а у некоторых рыб двойной) пары рёбер. Сегментами сформирована коробка для внутренних органов. Плавникообразиые грудные конечности располагаются непосредственно около головы, а тазовые конечности, за редким исключением, лежат у заднего конца туловища и не связаны посредством костей с осевым скелетом. Позади них помещается хвостовый отдел. С появлением в период эволюции ногообразных конечностей, т. е. на стадии хвостатых амфибий (рис. 29), отдел хвоста при движении в воде продолжает
Рис. 29. Схема скелета примитивного наземного позвоночного. 1—череп; 2—шейный отдел; 3—грудобрюшной отдел; 4—крестцовый позвонок; б—хвостовый отдел; 6—плечевой пояс; 7—тазовый пояс; 8—грудная конечность; 9—грудная кость; 10—тазовая конечность. играть старую роль двигателя, но при передвижении по дну водоёмов и на суше эту роль принимают на себя уже обе пары ногообразных конечностей. Главное усилие по толканию туловища вперёд падает на тазовые конечности. Поясок последних тесно соединяется с сегментами туловища, создавая более надёжную опору этим конечностям. С этого момента сегменты туловища, связывающиеся с поясом тазовых конечностей, становятся особым отделом, так называемым крестцовым, или, просто, крестц о м (4), правда, очень элементарным на первых порах, так как опирание происходит лишь на один сегмент. Он располагается на границе собственно туловищных сегментов и хвостовых. В начальном отделе собственно туловища, т. е. около головы, вырисовывается элементарный отдел шеи (2), так как первые от головы сегменты лишены полно развитых рёбер и конечности несколько отступают назад. С развитием ногообразных грудных конечностей связано также и появление элементарной грудной кости (9), в которую упирался пояс этих конечностей. У современных амфибий рёбра сильно укорочены. В дальнейшем усложнение скелета туловища в сторону более резких обособлений намеченных выше отделов осуществлялось уже у четвероногих предков млекопитающих, окончательно поселившихся на суше, т. е. близких к некоторым ископаемым рептилиям. В крестцовом отделе число сегментов увеличивается до двух и даже трёх. Кроме крестцового и хвостового отделов, на туловище уже резко обособляется шейный отдел, устройство которого давало большую свободу движению головы на туловище. Благодаря подвижности шейных сегментов голова может двигаться в разные стороны со значительной амплитудой и тем самым осуществляется возможность совершеннее и быстрее ориентироваться своими органами чувств в окружающей среде. Первая пара рёбер, замкнувшихся с вентральной стороны соответствующим куском грудной кости в полное кольцо, стала служить задней границей шейного отдела. Шейные рёбра недоразвиваются, и остатки их срастаются с соответствующими шейными позвонками. Внутренние органы шейного отдела туловища, сохранившиеся здесь в виде простых, недиференцирован-ных трубок—пищевода и воздухопровода (гортани и трахеи), становятся длиннее. В результате рельефного выделения крестца и шеи определённее обрисовался и промежуточный менаду ними отдел, который своим положением занимает, ориентировочно говоря, место между грудными и тазовыми конечностями. Этот участок (скелет туловища в узком смысле слова) является самым объёмистым и содержит в полостях тела всю главную массу диференцировавшихся внутренних органов. Он в свою очередь подразделяется на два отдела—грудной и поясничный (или брюшной). Характерным признаком грудного отдела служит наличие сильно развитых и подвижных в каждом сегменте рёбер, которые вместе с позвонками и грудной костью формируют грудную клетку как защитную коробку для органов дыхания, обособившихся от пищеварительной трубки. Кроме того, эта коробка обеспечивает покойную работу такого важного и нежного органа, как сердце. В поясничном отделе вентральные дуги, т. е. рёбра сегментов, укорачиваются, перестают быть подвижными, прочно срастаясь в виде рудиментов с позвонками. Вместе с позвонками они служат опорой и костной крышей брюшной полости, что особенно рельефно выражено у млекопитающих (рис. 39—7-? ). Мягкие, бесскелетные, т. е. лишённые рёбер, стенки этой полости ввиду своей податливости позволяют увеличивать ёмкость пищеварительной трубки до значительных размеров. Этим самым создаются условия для накапливания питательного материала, который транзитом и небольшими порциями проходит шейный и грудной отделы по пищеводу в пищеварительные органы брюшной полости, где и задерживается, подвергаясь переработке. Кроме того, рудиментарные ребра поясницы, сросшиеся с позвонками, служат и рычагами движения позвоночного столба. В туловище и хвосте довольно выпукло проявлено сегментальное расположение скелетных частей, хотя сегменты эти (позвонки), у позвоночных, строго говоря, являются вторичными, так как они развиваются в промежутках между сегментами первоначальной (примитивной) мускульной сегментации (миомерии). Чтобы разобраться в скелете туловища, необходимо предварительно ознакомиться с общими чертами построения костного сегмента и его элементами. Популярное:
|
Последнее изменение этой страницы: 2016-04-09; Просмотров: 1123; Нарушение авторского права страницы