|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Проявление свойств наследственного материала
На геномном уровне его организации
Самовоспроизведение и поддержание постоянства кариотипа в ряду поколений клеток
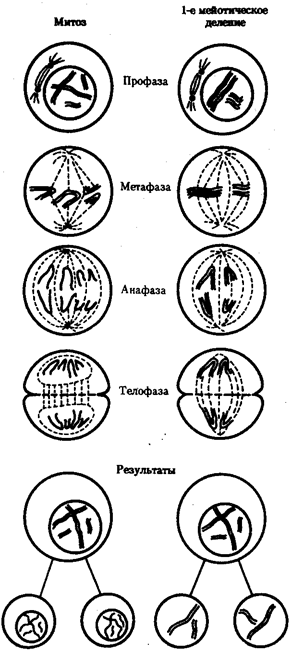
В основе самовоспроизведения кариотипа в ряду клеточных поколений лежат процессы самоудвоения отдельных хромосом, описанные в разд. 3.5.3.1. Жизнедеятельность клеток как структурно-функциональных единиц живого обеспечивается всей совокупностью получаемого ими генетического материала, поэтому особое значение приобретает поддержание постоянства кариотипа в ряду их поколений. На геномном уровне организации наследственного материала это обеспечивается всей совокупностью процессов, происходящих в митотическом цикле (см. гл. 2).
Рис. 3.68. Образование двуядерных клеток в результате торможения цитотомии при их делении Если содержание ДНК в гаплоидном наборе хромосом, т.е. в геноме, является видоспецифическим и соответствует с, то в диплоидном кариотипе, равном 2n, оно составляет 2с. В ходе митотического цикла количество ДНК в клетке изменяется от 2с (сразу после ее образования, когда хромосомы состоят из одной нити) до 4с (после репликации ДНК в S-периоде интерфазы, когда хромосомы становятся двунитчатыми). Число хромосом при этом остается неизменным и равным 2n на протяжении всего митотического цикла. Благодаря описанным ранее процессам, происходящим в ходе интерфазы и последующего митоза (см. гл. 2), дочерние клетки получают от материнской полноценную наследственную программу — генотип, заключенный в кариотипе. Однако не всегда клетки, удвоившие свой наследственный материал, вступают в митоз. Этим обусловлено существование в тканях некоторого количества клеток с формулой наследственного материала 2я4с, представляющих определенный резерв, готовый немедленно приступить к делению. Наконец, не всегда начавшееся деление заканчивается образованием двух дочерних клеток. Нарушение деления цитоплазмы при нормальном формировании дочерних ядер приводит к образованию двуядерных клеток (рис. 3.68). Разрушение ахромата-нового веретена, делающее невозможным распределение дочерних хромосом между его полюсами, сопровождается появлением полиплоидных клеток (рис. 3.69), в которых 4п дочерних хромосом остаются в одном неразделившемся ядре. Указанные отклонения от обычного течения митотического цикла являются причиной появления в тканях клеток, несущих больший по объему наследственный материал.
Рис. 3.69. Образование полиплоидньк клеток в результате разрушения веретена деления. А — различные наборы хромосом (12, 24, 48) в пыльцевых зернах одного из сортов гиацинта; Б — образование полиплоидных клеток /—в норме, II—при разрушении веретена деления колхицином Механизмы поддержания постоянства кариотипа В ряду поколений организмов
У организмов, размножающихся бесполым путем, новое поколение появляется из неспециализированных в отношении генеративной функции клеток тела. В основе их самовоспроизведения лежит митоз, обеспечивающий таким образом сохранение постоянной структуры наследственного материала в ряду поколений не только клеток, но и организмов.
При половом размножении процесс воспроизведения организмов осуществляется с участием специализированных половых клеток — гамет, вступающих в оплодотворение. При оплодотворении наследственный материал двух родительских гамет сливается, образуя генотип организма нового поколения — зиготы. Чтобы потомки получили соответствующую программу для развития видовых и индивидуальных характеристик, они должны обладать кариотипом, которым располагало предыдущее поколение. В такой ситуации поддержание постоянства кариотипа в ряду поколений организмов достигается предварительным уменьшением вдвое набора хромосом в гаметах, который восстанавливается до диплоидного при их оплодотворении: п + п = 2n. Образование гаплоидных гамет осуществляется в ходе гаметогенеза путем особой формы клеточного деления — мейоза. При мейозе из клеток с диплоидным набором In образуются гаметы с гаплоидным набором хромосом п (см. гл. 5). Такой результат достигается благодаря тому, что после однократного удвоения ДНК клетка делится дважды. В отличие от митоза в первом мейотическом делении в результате конъюгации гомологичные хромосомы объединяются в пары — биваленты. Последующее расхождение гомологов к разным полюсам веретена деления приводит к образованию клеток с гаплоидным набором хромосом: 2n4с → п2с. На рис. 3.70 представлены особенности первого деления мейоза в сравнении с митозом. В ходе второго мейотического деления сестринские хроматиды каждой хромосомы, как и в митозе, распределяются между дочерними клетками с наследственным материалом пс (рис. 3.71).
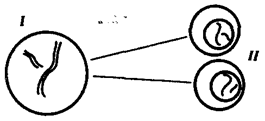
Рис. 3.71. Схема второго (эквационного) деления мейоза:
I — клетка, образованная в результате первого мейотического деления и несущая гаплоидный набор двунитчатых хромосом (n2c); II — клетки, образующиеся после второго деления мейоза и несущие гаплоидный набор однонитчатых хромосом (nc)
Благодаря особенностям мейоза образуются клетки, несущие полноценный геном, в котором каждая группа сцепления представлена в единственном экземпляре (гаплоидный набор хромосом). При самооплодотворении гаметы одного и того же родителя, а при перекрестном оплодотворении половые клетки разных организмов взаимодействуют друг с другом. Сперматозоиды, проникая в яйцеклетку, вводят в нее свой ядерный наследственный материал, заключенный в гаплоидном наборе хромосом. Ядра гамет сливаются и формируют диплоидное ядро зиготы, в котором каждая группа сцепления представлена в двойном экземпляре — отцовской и материнской хромосомами. Таким образом, мейоз и последующее оплодотворение обеспечивают сохранение у нового поколения организмов диплоидного кариотипа, присущего всем особям данного вида.
Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 686; Нарушение авторского права страницы