|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Взаимодействия между генами в генотипе
Отрицательные последствия нарушения дозового баланса связаны с тем, что генотип представляет собой не простую сумму отдельных генов. Гены в генотипе объединены в систему благодаря сложным и разнообразным взаимодействиям между ними, которые играют немаловажную роль в реализации информации, заключенной в каждом отдельном гене. Взаимодействие аллельных генов. Постоянно возникающие разнообразные изменения структуры генов обусловливают явление множественного аллелизма (см. разд. 3.4.2.4), поэтому взаимодействующие при оплодотворении гаметы часто несут в своих геномах разные аллели одного и того же гена. В связи с этим генотип нового организма является гетерозиготным по многим локусам, т.е. его аллельные гены, расположенные в соответствующих участках гомологичных хромосом и пришедшие от обоих родителей, представлены разными аллелями — А и А', В и В' С и С' и т.д. Если аллельные гены представлены одинаковыми аллелями, т.е. находятся в гомозиготном состоянии (АА или A'A', BB или B'B', CC или С'С'), то развивается соответствующий данному аллелю вариант признака. В случае гетерозиготности (АА', BB', CC') развитие данного признака (А, В или С) будет зависеть от взаимодействия аллельных генов. Доминирование — это такое взаимодействие аллельных генов, при котором проявление одного из аллелей (А) не зависит от присутствия в генотипе другого аллеля (А') и гетерозиготы АА' фенотипически не отличаются от гомозигот по этому аллелю (АА). Такая ситуация наблюдается, например, когда один из аллелей гена А («дикий») способен обеспечить формирование определенного варианта признака (синтез пептида с определенными свойствами), а другой А'—не обладает такой способностью. Наличие в генотипе АА' единственного нормального аллеля А приводит к формированию нормального признака. Этот аллель выступает как доминантный в данном гетерозиготном генотипе. Присутствие другого аллеля (А') фенотипически не проявляется, поэтому его называют рецессивным. Примером доминирования одного из алледей в гетерозиготном генотипе может служить определение групповой принадлежности крови у человека по системе АВ0. Генотипы, содержащие аллель IA либо в гомозиготном состоянии, либо в сочетании с аллелем I0 (IAIA или IAI0), определяют развитие у человека второй группы крови (группа крови А). Такая же ситуация наблюдается и в отношении аллеля IB, обусловливающего формирование третьей, или В-группы крови. Следовательно, аллели IA и IB выступают как доминантные по отношению к аллелю I0, формирующему в гомозиготном состоянии I0I0 первую, или 0-группу крови. Неполное доминирование наблюдается, когда фенотип гетерозигот BB' отличается от фенотипа гомозигот по обоим аллелям (BB или B'B') промежуточным проявлением признака. Это объясняется тем, что аллель, способный сформировать нормальный признак, находясь в двойной дозе у гомозиготы BB, проявляется сильнее, чем в единственной дозе у гетерозиготы BB'. Указанные генотипы отличаются экспрессивностью, т.е. степенью выраженности признака. Демонстрацией такого типа взаимодействия генов могут быть многочисленные наследственные заболевания у человека, проявляющиеся клинически у гетерозигот по мутантным аллелям, а у гомозигот заканчивающиеся смертью. Иногда гетерозиготы имеют почти нормальный фенотип, а гомозиготы характеризуются пониженной жизнеспособностью. Так, гомозиготы по аллелю серповидноклеточности эритроцитов в связи с развитием у них тяжелой формы анемии и других фенотипических проявлений (см. рис. 3.21) обычно не переживают детский возраст. Напротив, гетерозиготы — это как правило нормальные люди. Вместе с тем, они все-таки испытывают кислородную недостаточность в большей степени в сравнении с гомозиготами по аллелю дикого типа, в частности при подъеме на высоту (см. 4.1). Кодоминирование представляет собой такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате этого формируется некий промежуточный вариант признака, новый по сравнению с вариантами, определяемыми каждым аллелем самостоятельно. Примером может служить формирование IV, или АВ-группы, крови у человека, гетерозиготного по аллелям IA и IB, которые по отдельности детерминируют образование II и III групп крови. Межаллельная комплементация относится к достаточно редко встречаемым способам взаимодействия аллельных генов. В этом случае возможно формирование нормального признака D у организма, гетерозиготного по двум мутантным аллелям гена D(D'D" ). Допустим, что ген D отвечает за синтез какого-то белка, который имеет четвертичную структуру, состоящую из нескольких одинаковых пептидных цепей. Мутантный аллель D' определяет синтез измененного пептида D', a мутантный аллель D" приводит к синтезу другой, но тоже измененной структуры пептида D". Можно представить ситуацию, когда взаимодействие таких измененных пептидов (D' и D" ) при формировании четвертичной структуры, как бы взаимно компенсируя эти изменения, обеспечивает образование белка с нормальными свойствами. В то же время отдельно взаимодействующие пептиды D' или D" формируют аномальные белки. Таким образом, с определенной вероятностью у гетерозигот D'D" в результате межаллельной комплементации может образовываться нормальный признак в виде белка с нормальными свойствами. Аллельное исключение—такой вид взаимодействия аллельных генов в генотипе организма, который можно понять на примере рассмотренного выше механизма инактивации одной из Х-хромосом у особей гомогаметного пола, приводящего в соответствие дозы Х-генов у всех представителей вида. Инактива-ция одного из аллелей в составе Х-хромосомы способствует тому, что в разных клетках организма, мозаичных по функционирующей хромосоме, фенотипически проявляются разные аллели (рис. 3.79). Аллельное исключение наблюдается также в В-лимфоцитах, синтезирующих специфичные антитела к определенным антигенам. Моноспецифичность таких иммуноглобулинов требует выбора, который должна осуществить каждая клетка между экспрессией отцовского или материнского аллеля.
Рис. 3.79. Мозаицизм женского организма по наличию или отсутствию нормальных потовых желез в коже, обусловленный экспрессией нормального или мутантного аллелей гена Х-хромосомы
Затемнены участки кожи, лишенные потовых желез, в клетках которых экспрессируется мутантный аллель Таким образом, даже процесс формирования элементарного признака — синтез полипептида с определенной последовательностью аминокислот — зависит, как правило, от взаимодействия по меньшей мере двух аллельных генов, и конечный результат определяется конкретным сочетанием их в генотипе. Формирование сложных признаков предполагает необходимость взаимодействия неаллельных генов, занимающих разное положение в геноме данного вида. Взаимодействие неаллельных генов. Большинство признаков и свойств организма, по которым он отличается от других представителей вида, являются результатом действия не одной пары аллельных генов, а нескольких неаллельных генов или их продуктов. Поэтому эти признаки называют сложными. Сложным признаком может быть четвертичная структура белка, образуемая разными пептидными цепями, которые детерминируются разными (неаллельными) генами. Например, молекула гемоглобина, содержит две о, - и две (3-цепи, гены которых расположены у человека в 16-й и 11-й хромосомах (см. разд. 3.6.4.3). Сложный признак может быть обусловлен совместным однозначным действием нескольких генов или являться конечным результатом цепи биохимических преобразований, в которых принимают участие продукты многих генов. Наконец, на формирование как простых, так и сложных признаков определенные регуляторные воздействия оказывают другие локусы. В зависимости от характера участия неаллельных генов в формировании того или иного признака различают несколько видов их взаимодействия. Большинство количественных признаков организмов определяется полигенами, т.е. системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов в процессе формирования признака называют полимерным. Оно сводится чаще всего к суммированию действия сходных аллелей этих генов, определяющих формирование одинакового варианта признака. Совместное действие полигенов обусловливает различную экспрессивность — степень выраженности признака, зависящую от дозы соответствующих аллелей. В основе появления в геноме таких генов, очевидно, лежит их дупликация или амплификация (см. разд. 3.6.4.3). Это позволяет увеличить синтез соответствующего продукта в клетках организма. По полимерному типу взаимодействия у человека определяется интенсивность окраски кожных покровов, зависящая от уровня отложения в клетках пигмента меланина. В геноме человека имеется четыре гена, отвечающих за этот признак. В генотипе все они представлены в двойной дозе. В связи с тем что эти гены отвечают за развитие одного и того же признака, их обозначают одной и той же буквой, но с разными символами, чтобы подчеркнуть их неаллельность — P1P2P3P4. Наличие в генотипе восьми доминантных аллелей в системе полигенов, определяющих цвет кожи, обусловливает максимальную ее пигментацию, наблюдаемую у африканских негров (P1P1P2P2P3P3P4P4). Полное отсутствие доминантных аллелей у. рецессивных гомозигот (р1р2р2р3р3р4р4) проявляется в виде минимальной пигментации у европеоидов. Большее или меньшее количество доминантных аллелей, колеблющееся от 8 до 0, обеспечивает разную интенсивность окраски кожи (рис. 3.80). Полимерное взаимодействие генов лежит в основе определения главным образом количественных признаков (рост, масса организма, возможно, интеллект).
Рис. 3.80. Зависимость интенсивности пигментации кожи у человека от количества доминантных аллелей в системе полигенов (Р) в генотипе
Остальные пояснения см. в тексте
Большинство сложных признаков, являясь результатом целой цепи биохимических и структурных преобразований, требует участия многих генов, влияющих на разные звенья этого процесса. Отсутствие нормального первичного продукта, хотя бы одного из них, не позволяет сформировать нормальный сложный признак. Так как часто нормальный первичный продукт гена является результатом экспрессии его доминантного аллеля, сложный признак формируется при наличии в генотипе доминантных аллелей всех взаимодействующих генов. Такое взаимодействие неаллельных генов, при котором они взаимно дополняют друг друга, называют комплементарным взаимодействием. В качестве примера комплементарного взаимодействия неаллельных генов можно рассмотреть процесс формирования половой принадлежности организма у человека. Развитие признаков пола у человека, так же как у большинства животных, определяется в первую очередь сочетанием гетерохромосом в его кариотипе. Наличие Х- и Y-хромосом обусловливает возможность формирования мужского пола, а двух Х-хромосом —женского. Однако установлено, что для развития организма мужского пола не достаточно присутствия одного лишь Y-сцепленного гена, который определяет дифференцировку половых желез по мужскому типу и синтез ими гормона тестостерона. Для этого необходим также продукт другого гена — белок-рецептор, обеспечивающий проникновение гормона в клетки тканей-мишеней. За синтез такого белка отвечает особый ген, расположенный в Х-хромосоме. Его мутация, нарушающая образование нормального белка-рецептора, делает ткани-мишени невосприимчивыми к гормону, направляющему их развитие по мужскому типу. Не использовав такую возможность на определенном этапе онтогенеза, организм осуществляет развитие по женскому типу. В результате появляется особь с кариотипом XY, но внешне более сходная с женщиной. Такие субъекты не способны иметь потомство, так как их половые железы (семенники) недоразвиты, а их выводные протоки часто формируются по женскому типу (недоразвитая матка, влагалище). Вторичные половые признаки также характерны для женского пола. Описанная картина известна у человека как тестикулярная феминизация, или синдром Морриса (рис. 3.81).
Рис. 3.81. Тестикулярная феминизация —синдром Морриса: I — кариотип XY (удаление семенников в детском возрасте); II — евнухоидная форма, кариотип XY (отсутствие молочных желез, вторичного оволосения, естественного влагалища); III — кариотип XY
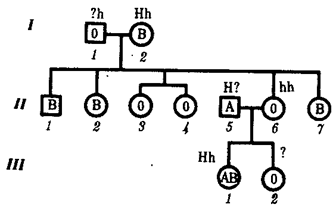
Таким образом, достижение конечного результата при формировании у человека признаков мужского пола является следствием взаимодополняющего действия нескольких (по меньшей мере двух) генов, определяющих возможность синтеза мужского полового гормона и белка-рецептора. Другим примером взаимодействия неаллельных генов служит формирование у человека групповой принадлежности крови по системе АВО. Выше этот признак рассматривался с точки зрения взаимодействия аллельных генов. Отвечающий за формирование данного признака ген I обеспечивает синтез антигенов А и В, фиксирующихся на поверхности эритроцитов. Однако для синтеза антигенов А и В необходимо образование неспецифического вещества-предшественника, которое детерминируется геном Н, находящимся в доминантном состоянии. У гомозигот по рецессивному аллелю hh даже при наличии в генотипе доминантных аллелей гена ^ или Iй антигены А и В не синтезируются и фенотипически эти люди имеют I группу крови.
Рис. 3.82. Родословная, иллюстрирующая наследование группы крови по системе АВО при бомбейском феномене
Женщина II.6 с группой крови 0(1) имеет ребенка III.1 с группой крови АВ (ГУ); вероятно, в ее геноме имеется аллель Iе, полученный ею от матери (1.2), но не проявившийся в связи с ее гомозиготностыо по рецессивному аллелю hh. Бомбейский фенотип встречается с частотой 1: 13000 среди индусов, живущих в окрестностях Бомбея Такая ситуация характерна для бомбейского феномена, описанного впервые в семье, где у женщины I группы крови родился ребенок с IV группой крови. Очевидно, ее фенотип был обусловлен непроявлением имеющегося в генотипе аллеля IВ, что связано с гомозиготностью матери по рецессивному аллелю hh (рис. 3.82). Вероятно с этих же позиций можно рассматривать и случаи альбинизма у человека, когда даже у африканских негров, которые обладают максимумом доминантных аллелей в системе полигенов, определяющих пигментацию кожи, появляются альбиносы с полным отсутствием пигмента в клетках кожи, радужке глаз, волосах (рис. 3.83). Альбинизм связывают с гомозиготностыо генотипа по рецессивному аллелю гена, не относящегося к полигенам пигментации. Возможно, его доминантный аллель определяет синтез продукта, принимающего участие в процессах, предшествующих синтезу пигмента. В таком случае здесь также имеет место взаимодополняющее действие соответствующих аллелей разных генов. В некоторых случаях при взаимодействии неаллельных генов для развития сложного признака необходимо обязательное присутствие одного из генов (А) в гомозиготном рецессивном состоянии (аа), тогда другой ген (В) обеспечивает формирование признака. Наличие в генотипе доминантного аллеля гена А каким-то образом препятствует проявлению гена В(b), и признак не формируется. Такое взаимодействие неаллельных генов принято называть эпистатическим,
Рис. 3.83. Пример рождения альбиноса у негритянки (отсутствие пигментации кожи у ребенка может быть результатом рецессивного эпистаза)
Примером эпистатического взаимодействия генов может быть подавление у многих видов развития окраски покровов, определяемое одним из генов, при наличии в генотипе другого гена в доминантном состоянии. Так, у тыквы развитие окраски плодов определяется геном В. Доминантный его аллель детерминирует желтую, а рецессивный — зеленую окраску. Однако окраска не развивается вообще, если в генотипе имеется неаллельный ген А в доминантном состоянии. У кур породы леггорн белое оперение является результатом эпистатического воздействия доминантного аллеля одного гена на проявление неаллельного ему гена, отвечающего за пигментацию оперения. Отсутствие необходимых данных о роли первичных продуктов многих генов в формировании сложных признаков часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образования этих признаков. В одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное взаимодействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантном состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантном эпистазе. Вероятно, само разделение взаимодействия генов на комплементарное и эпистатическое несколько искусственно, ибо во всех этих случаях сложный признак является результатом сочетания в генотипе определенных аллелей соответствующих генов, которые обеспечивают синтез продуктов, участвующих в цепи биохимических преобразований на разных уровнях формирования сложного признака. Так как при определенных сочетаниях аллелей неаллельных генов становится невозможным фенотипическое проявление доминантных аллелей некоторых из них, нередко наблюдается неполная пенетрантность доминантных аллелей — они проявляются не у всех носителей. Так, аллели IA и IB, определяющие групповую принадлежность крови по системе АВО, не проявляются фенотипически в отсутствие доминантного аллеля гена Н (бомбейский феномен). У альбиносов (аа) не проявляются доминантные аллели полигенов пигментации кожных покровов (Р1Р2Р3Р4). У тыквы не развивается окраска плодов, а у кур породы леггорн — пестрая окраска оперения, контролируемые доминантными аллелями соответствующих генов, при наличии в их генотипе определенных неаллельных генов в доминантном состоянии. Особый вид представляет взаимодействие, обусловленное местом положения гена в системе генотипа, — эффект положения. Непосредственное окружение, в котором находится ген, может сказываться на характере его экспрессии. Изменение активности гена, наблюдаемое при хромосомных перестройках, нередко связано с перемещением его в другую группу сцепления при транслокациях или изменением его положения в своей хромосоме при инверсиях. Особый случай, очевидно, представляет изменение экспрессии генов в результате деятельности подвижных генетических элементов, активирующих или угнетающих проявление генов, вблизи которых они встраиваются. Наконец, большое значение в объединении генов в единую систему генотипа имеют регуляторные взаимодействия, обеспечивающие регуляцию генной активности. Продукты генов-регуляторов — белки-регуляторы — обладают способностью узнавать определенные последовательности ДНК, соединяться с ними, обеспечивая, таким образом, транскрибирование информации со структурных генов или препятствуя транскрипции (см. разд. 3.6.6).
Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 747; Нарушение авторского права страницы