|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Коры у ребенка по отношению к поверхности полушария
взрослого (по возрастам), %
Аналогичным образом развиваются в онтогенезе верхние (ассоциативные) слои только что упомянутых зон коры. И здесь мы отмечаем уже знакомый нам факт особенно интенсивного роста ширины этих функционально наиболее важных слоев коры к 3 — 3, 5 года жизни ребенка, причем увеличение их в случае некоторых особенно сложных полей продолжается до 7- и даже 12-летнего возраста. Этот факт отчетливо говорит о том, что по мере развития ребенка возрастает роль тех видов деятельности, которые требуют совместной работы отдельных зон коры и осуществляются при ближайшем участии верхних, ассоциативных, или интегративных, ее слоев.
Более подробное описание только что указанной тенденции содержится в таблице 6, составленной на основании работ целой группы отечественных морфологов. Из таблицы видно, что если для наиболее простых (первичных, или проекционных) зон коры характерно незначительное развитие в онтогенезе, то более сложные — вторичные и третичные — зоны коры развиваются в онтогенезе весьма интенсивно.
Увеличение площади вторичных и третичных зон коры на сравнительно поздних этапах филогенеза является далеко не единственным признаком, позволяющим судить о их готовности к участию в регуляции поведения человека. Столь же существенной характеристикой является степень миелинизации соответствующих нервных образований.
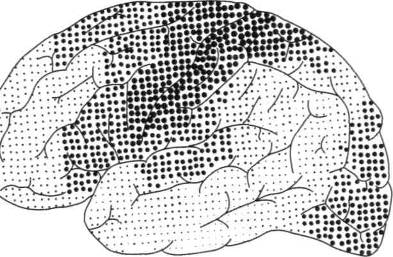
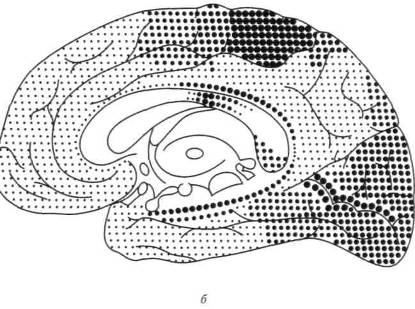
Как показали классические исследования Флексига (1900, 1927), процесс миелинизации — по завершении которого нервные элементы становятся готовыми к нормальному функционированию — протекает в разных зонах коры неравномерно: если миелинизация элементов первичных (проекционных) зон заканчивается довольно рано, то процесс миелинизации во вторичных и третичных зонах коры затягивается на очень длительные сроки и в некоторых случаях продолжается до 7 —12-летнего возраста. Достаточно посмотреть на миелогенетическую карту, составленную Фогтами (рис. 21), на которой изображены зоны, элементы которых наиболее рано кончают миелинизацию (крупные кружки), зоны, которые миелинизируются в последнюю очередь (мелкие точки), и зоны, занимающие промежуточное место (средние точки), чтобы отчетливо увидеть, что аппараты, соответствующие наиболее сложным, комплексным формам психической деятельности созревают на относительно поздних этапах развития и что, следовательно, формирование психической деятельности человека идет от более простых к сложным, опосредствованным формам.
Рис. 21. Миелогенетическая карта Фогтов. Крупными точками обозначены наиболее рано созревающие отделы коры. Мелкими точками — наиболее поздно созревающие отделы коры: а — наружная поверхность; 6 — внутренняя поверхность
Далее, на протяжении книги, нам не один раз представится случай убедиться в том, насколько большое значение имеют полученные в эволюционной анатомии данные для решения важнейших проблем психологической науки.
2 ФИЗИОЛОГИЧЕСКИЕ ДАННЫЕ: МЕТОД РАЗДРАЖЕНИЯ
Опыты с непосредственным раздражением коры
Данные, полученные в сравнительно-анатомических исследованиях, составляют основу наших знаний о функциональной организации мозга как органа психической жизни; однако они не дают прямых указаний на нервные механизмы, связанные с приемом информации от внешнего мира и регуляцией поведения человека.
Существенный прогресс в этом направлении связан с применением метода раздражения отдельных участков мозга. Благодаря ему физиологи смогли получить сведения о непосредственной функции тех или иных мозговых систем.
Еще во второй половине XIX века исследователи раздражали отдельные пункты коры головного мозга животных, с тем чтобы выявить таким путем функции различных участков мозга. В 1871 г. были опубликованы результаты известных опытов Фрича и Гитцига, установивших, что раздражение электрическим током определенных участков коры головного мозга собаки вызывает сокращение мышц противоположных конечностей. Так была впервые выделена двигательная зона коры и положено начало точному физиологическому исследованию мозговых функций (рис. 22).
Рис. 22. Двигательные и сенсорные реакции при раздражении пре- и постцентральных отделов коры (по Пенфилду и Расмуссену): а — представительство двигательных процессов в мозговой коре; б — представительство сенсорных процессов в мозговой коре. Длина столбиков обозначает относительное число моторных и сенсорных реакций, вызванных раздражением точек, расположенных спереди и сзади от роландовой борозды
Спустя сравнительно небольшой период времени классические исследования, проведенные Чарльзом Шеррингтоном и его сотрудниками (Грюнбаум и Шеррингтон, 1903; Лейтон и Шеррингтон, 1917), показали, что двигательная зона коры головного мозга обезьяны имеет четкую функциональную организацию. Оказалось, что гигантские пирамидные клетки верхних отделов передней центральной извилины дают начало волокнам, идущим к нижним, а гигантские пирамидные клетки ее нижних отделов — к волокнам, идущим к верхним конечностям противоположной стороны тела. (Раздражение соответствующих пунктов двигательной зоны слабым электрическим током вызывало сокращение строго определенных мышц противоположной стороны тела.) Опыты Шеррингтона и его сотрудников положили начало изучению функциональной организации мозговой коры объективными физиологическими методами. Успехи нейрохирургии очень скоро сделали возможным дальнейший прогресс в этой области; через два десятилетия после первых публикаций Шеррингтона О. Ферстер, один из основоположников нейрохирургии, смог показать, что предложенные Шеррингтоном методические принципы пригодны и для анализа функций коры головного мозга человека. Последовавшие за исследованиями О. Ферстера работы Г. Хоффа и О.Петцля (1930) и, наконец, выдающегося канадского нейрохирурга В. Пенфилда и его сотрудников (Пенфилд и Эриксон, 1945; Пенфилд и Расмуссен, 1950; Пенфилд и Джаспер, 1959) создали прочную основу учения о функциональной организации различных зон мозговой коры человека.
Опыты Пенфилда не только подтвердили, что сформулированный Шеррингтоном применительно к мозгу обезьяны принцип функциональной организации моторной зоны коры верен и для мозга человека, в них были разработаны два принципиально новых аспекта исследования.
Во-первых, возможность наблюдать человека во время тонких нейрохирургических операций позволила расширить сферу изучения от двигательных функций коры до ее сенсорных функций: раздражая кору задней центральной извилины, а также затылочной и височной областей, исследователь мог спрашивать у больного об испытываемых им ощущениях и таким образом выявлять сенсорные функции этих зон коры. Таким образом, очень скоро оказалось возможным, не ограничиваясь исследованием функциональной организации двигательной сферы мозга, перейти к изучению функциональной организации афферентных, сенсорных систем.
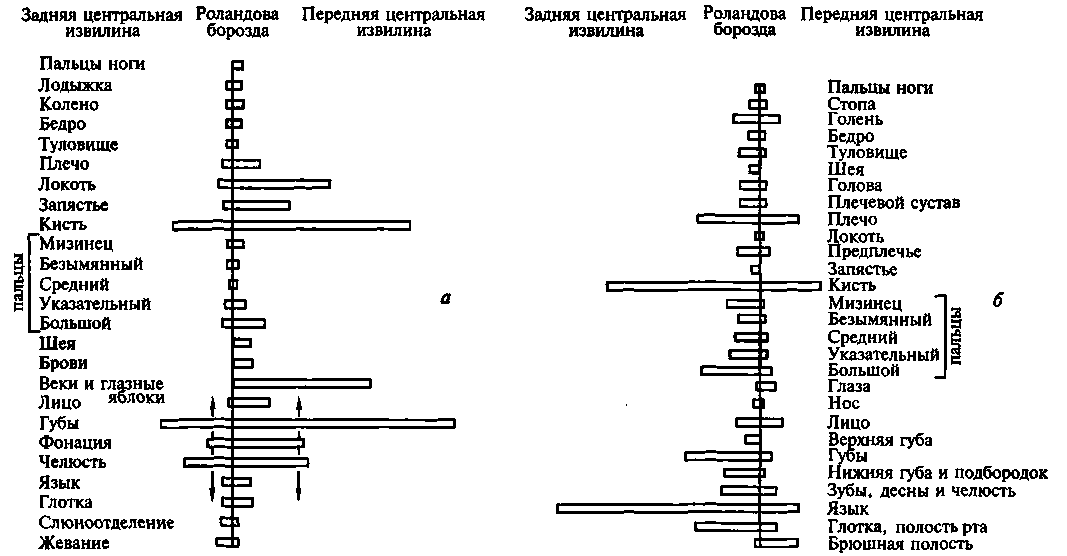
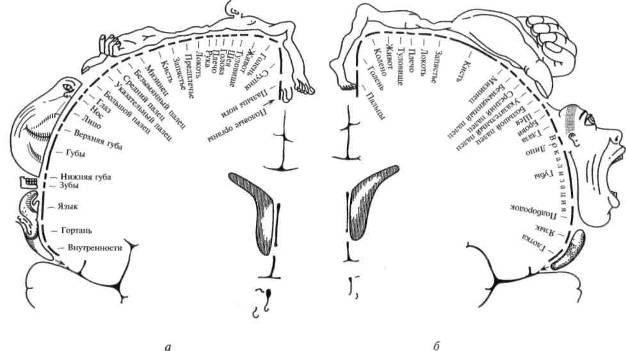
Во-вторых, эти исследования позволили выйти за пределы простой констатации соответствия отдельных пунктов коры определенным участкам тела или, как это принято было говорить, принципа соматотопической организации первичных зон мозговой коры. Пенфилд показал, что как передняя, так и задняя центральные извилины организованы не столько по принципу линейной зависимости между размерами соответствующих участков тела и их проекциями на кору головного мозга, сколько по функциональному принципу: чем большее значение имеет та или другая функциональная система, тем более обширную территорию занимает ее проекция в первичных отделах коры 'головного мозга.
Оказалось, что проекции туловища, бедра или плеча занимают в передней и задней центральных извилинах относительно небольшую площадь; проекция руки — гораздо большую, а проекции губ и языка — еще большую площадь (рис. 23). Оказалось далее, что проекции 3-го и 4-го пальцев в коре сравнительно малы, тогда как участки, раздражая которые можно вызвать движения большого или указательного пальца, очень велики.
Рис. 23. Схема проекционных путей первичных моторных и сенсорных зон коры (по Пфейфферу и Кригу)
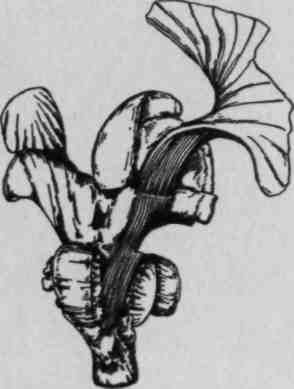
Так возникли известные схемы Пенфилда (рис. 24), наглядно показывающие, что чем более управляемым является тот или иной орган, тем шире он представлен в мозговой коре.
Рис. 24. Сравнительная площадь проекции отдельных поверхностей тела в проекционных зонах мозговой коры (по Пенфилду): а — корковая проекция чувствительности; 6 — корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения
Еще более углубили наши представления о физиологических механизмах первичных зон коры исследования, в которых было показано, что колонки клеточных элементов, соответствующие функционально наиболее важным системам органов, наиболее чувствительны к раздражениям и что существует «дифференциальная реактивность» (Differential accessibility) систем нервного аппарата, имеющих различное функциональное значение (Маунт-кастл, 1966; Филлипс, 1956, 1966). В этом отношении важны также многочисленные данные, показывающие, что пороги возбудимости клеточных элементов могут меняться под влиянием воздействий со стороны нижележащих отделов мозга (Мэгун и Моруцци, 1949; Линдсли и др., 1949; Френч, 1952; Джаспер, 1963, 1966; Эрнандес-Пеон, 1966; и др.).
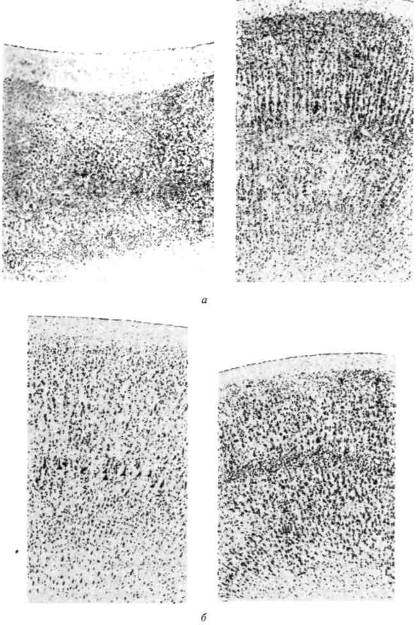
Применение описанных физиологических приемов исследования функциональной организации мозга не ограничилось опытами с раздражением первичных зон мозговой коры. Результаты, значение которых для понимания функциональной организации коры головного мозга трудно переоценить, были получены в экспериментах со стимуляцией вторичных отделов мозговой коры, отличающихся, как было указано ранее, мощным развитием верхних (ассоциативных) слоев (рис. 25).
Рис. 25. Цитоархитектоническое строение первичных и вторичных зон коры (по Бродману): а — задние отделы коры; б — передние отделы коры
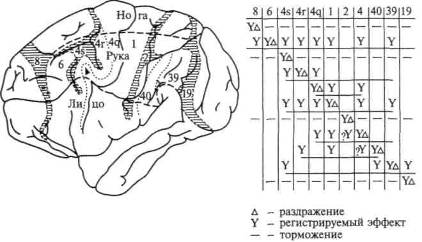
Едва ли не самые интересные результаты были получены американским исследователем Мак-Кэллоком (1943, 1944), сумевшим ответить на вопрос о том, как далеко распространяется возбуждение, вызываемое раздражением определенного пункта коры головного мозга. Предложенный автором нейронографический метод был очень прост: небольшую бумажку, смоченную раствором стрихнина, налагали на определенный участок коры головного мозга, тем самым раздражая его. Затем электроды последовательно прикладывали к соседним участкам, «прощупывая» таким образом, далеко ли распространяется вызванное раздражение.
Эксперимент показал, что если раздражение первичных зон коры распространялось лишь на области, непосредственно прилегающие к раздражаемому пункту, то возбуждение, вызванное раздражением вторичных зон коры, охватывало целый комплекс зон коры, иногда расположенных на значительном расстоянии от раздражаемого пункта (рис. 26). Этот факт свидетельствует о том, что в процесс возбуждения, возникающий во вторичных отделах коры, вовлекаются большие системы нервных элементов и, таким образом, обеспечиваются значительно более сложные, интегральные, процессы по сравнению с теми, которые имеют место при возбуждении первичных зон.
Рис. 26. Данные нейронографических опытов с раздражением первичных и вторичных (сенсорных и двигательных) отделов коры (по Мак-Кэллоку). Схема показывает, как далеко распространяется возбуждение, вызванное раздражением стрихнином определенного участка коры головного мозга шимпанзе. Поля 6, 4, 1 относятся к сенсомоторной коре. Заштрихованы так называемые депрессорные поля
Значение такого обширного распространения возбуждения, исходящего из вторичных зон мозга, становится ясным из опытов, в которых эффект раздражения проверялся с помощью не электрофизиологических, а психологических показателей. Результаты этих опытов настолько важны, что на них следует остановиться особо.
Эксперименты с раздражением коры головного мозга больного на операционном столе, проведенные О.Петцлем, а затем Пенфилдом и его сотрудниками, показали, что эффект раздражения определенных пунктов первичных зон коры резко отличен от того, который вызывается раздражением вторичных зон. Это с особенной отчетливостью видно при раздражениях соответствующих отделов затылочной (зрительной) и височной (слуховой) коры.
Раздражение первичных отделов коры вызывало у больного элементарные ощущения. Так, больной, у которого на операционном столе раздражали полюс затылочной зоны, заявлял, что он внезапно начинает видеть мелькающие световые точки, окрашенные шары, языки пламени и т.п., причем эти неоформленные зрительные галлюцинации (фотопсии) располагались в различных участках зрительного поля в строгой зависимости от того,
какое место коры раздражалось (они появлялись в верхних секторах поля зрения, если раздражались нижние участки зрительной коры, и в нижних секторах, если раздражались верхние участки). Аналогичная картина наблюдалась при раздражении первичных участков височной (слуховой) коры, с той только разницей, что в этих случаях у человека появлялись элементарные слуховые галлюцинации (шумы, тоны).
Совершенно иными явлениями сопровождается раздражение вторичных отделов коры. Раздражение передних отделов затылочной области (вторичной зрительной коры) вызывает сложные оформленные зрительные образы: испытуемый видит людей, зверей, бабочек, человека, идущего или делающего знаки рукой, птиц в полете и т. п. Следовательно, возбуждение, вызванное раздражением этой области коры, не только распространяется на более обширные площади, но и приводит к всплыванию законченных зрительных образов, сформировавшихся у человека в прежнем опыте.
Аналогичные факты отмечаются при раздражении соответствующих отделов височной (слуховой) коры. Если, как уже было указано, раздражение первичных отделов слуховой коры (трудноосуществимое из-за того, что первичные зоны слуховой коры расположены в малодоступной поперечной извилине височной области) вызывает лишь ощущение тонов или шумов, то раздражение вторичных отделов слуховой коры, расположенных в верхней и средней извилинах височной области, приводит к появлению сложных слуховых галлюцинаций — музыкальных мелодий, а иногда (если раздражается височная кора левого полушария) к звучанию слов, фраз, песен, которые испытуемый слышит, полностью сознавая отсутствие их внешнего источника, так что они оказываются чем-то средним между реальными слуховыми ощущениями и слуховыми воспоминаниями.
Интересны опыты с раздражением еще более сложных отделов височной коры. Пенфилд, который провел большую серию таких исследований, мог наблюдать, что раздражение этих областей коры вызывает сложные сценоподобные галлюцинации — целые картины, иногда включающие в свой состав как зрительные, так и звуковые компоненты. Помимо опытов с раздражением определенных пунктов коры электрическим током аналогичные явления наблюдаются в тех случаях, когда рубец, образовавшийся в мозговой ткани (или оболочке мозга) после ранения или кровоизлияния, оказывает постоянное раздражающее воздействие на соответствующие участки мозга. Накопление такого раздражения может привести к эпилептическим припадкам (например, к так называемой Джексоновской эпилепсии), причем этим припадкам предшествуют явления, носящие характер зрительных или слуховых предвестников (ауры), сложность которых зависит от локализа-
ции рубца. Если рубец расположен в пределах первичных зон зрительной или слуховой коры, аура носит характер элементарных зрительных или слуховых галлюцинаций; если рубец расположен в более сложных — вторичных областях коры, она может принимать характер сложных, оформленных (зрительных или слуховых) галлюцинаций. Таким образом, характер ауры, предшествующей эпилептическому припадку, служит для невропатолога указанием на местоположение рубца, являющееся «эпилептогенной зоной».
Дополнение представлений об анатомическом строении отдельных зон мозговой коры данными, полученными при раздражении ее различных участков, позволяет значительно расширить наше понимание функциональной организации коры человеческого мозга.
Обе эти группы фактов подтверждают сформулированное ранее положение о том, что отдельные системы коры головного мозга имеют иерархическое строение и что возбуждение, возникающее в периферических органах чувств, сначала приходит в первичные (проекционные) зоны, отдельные участки которых представляют собой расположенные по топографическому принципу проекции соответствующих периферических рецепторов, и лишь вслед за этим распространяется на вторичные зоны коры, которые, опираясь на аппарат верхних (ассоциативных) слоев нейронов, играют интегрирующую роль, объединяя топографические (соматотопические) проекции возникших на периферии возбуждений в сложные функционально организованные системы.
По иерархическому принципу построены и двигательные отделы коры: над проекционными отделами передней центральной извилины (моторная зона) надстраиваются вторичные отделы премоторной области, располагающие мощно развитыми верхними (ассоциативными) слоями нейронов и способные превратить отдельные двигательные импульсы в целые системы возбуждений, составляющие физиологическую основу сложных предметных движений и двигательных навыков. Подробно о функциональной организации систем двигательной коры мы будем говорить далее.
Открытие системного характера построения основных зон мозговой коры является одним из главнейших этапов формирования наших представлений о функциональной организации человеческого мозга, и мы еще много раз сможем убедиться в важности этого фундаментального принципа.
Опыты с непрямой стимуляцией коры
До сих пор мы останавливались на результатах классических исследований, основанных на непосредственном электрическом или механическом раздражении коры головного мозга.
Существуют и другие, гораздо более естественные методы изучения функций головного мозга, которые, однако, настолько давно известны в психологии и физиологии, что мы упомянем о них здесь лишь в самых общих чертах. Некоторые из этих методов легли в основу объективного изучения поведения животного. Животному, например, предъявляли какой-нибудь стимул (обычно связанный с инстинктивной деятельностью и являющийся для него безусловным раздражителем), после чего изучали изменения в поведении животного, которые вызывались этим стимулом. И.П.Павлов предложил метод условных рефлексов, где условные раздражители (свет, звук, прикосновение), сами по себе способные вызвать лишь элементарную ориентировочную реакцию, в сочетании с безусловными раздражителями вызывают специфические — пищевые, оборонительные или половые — реакции. Этот метод открыл новые перспективы в изучении мозговой деятельности.
Работы Павлова настолько известны, что мы не будем останавливаться на них специально. Один из приемов «непрямой» стимуляции мозговой коры заслуживает, однако, того, чтобы на нем остановиться особо. Заключается этот прием в том, что раздражение адресуется не непосредственно коре, а периферическому отделу анализатора (коже, глазу, уху), после чего в отдельных участках коры головного мозга или в подкорковых образованиях прослеживается эффект такого раздражения. Один из наиболее важных вариантов этого метода называется методом вызванных потенциалов.
Рассмотрим этот метод подробнее. Исследователи, применявшие метод раздражения в его наиболее простых формах, раздражали определенные пункты коры головного мозга электрическим током и описывали те изменения в поведении животных или человека, которые возникали как следствие такого раздражения.
Существует, однако, обратная возможность: если поставить животное или человека в условия, когда на него воздействуют определенные естественные факторы, можно выявить, какие участки коры реагируют на эти воздействия изменением электрической активности. Такой прием очень распространен в современной электрофизиологии и применяется для исследования функций различных участков коры больших полушарий.
Вариант этого метода, предложенный Даусоном (1947), получил особенно большое распространение и позволил с достаточной отчетливостью показать, как именно распространяется система импульсов, вызванных зрительными (Гранит, 1956), слуховыми (Эрнандес-Пеон, 1959, 1961) или кожными (Маунткастл, 1957, 1961, 1966; и др.) раздражителями, и детально изучить те влияния, которые оказывают нижележащие нервные образования ствола мозга на состояние мозговой коры (Эди, 1956; Эрнандес-Пеон, 1959, 1961; Линдсли, 1960, 1961).
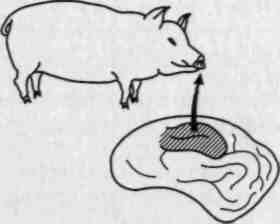
Наибольший интерес для учения о функциональной организации отдельных систем мозговой коры представляют, однако, данные Эдриана (1936), показавшего, что в отдельных зонах мозговой коры сенсорная периферия представлена неравномерно и что функционально наиболее важным системам в коре соответствуют большие площади. Так, оказалось, что раздражение бедра животного вызывает изменение потенциалов лишь в незначительных по величине участках коры головного мозги животного, в то время как раздражение функционально значимых органов, играющих особенно большую роль в его жизнедеятельности (например, пятачка у свиньи, губ у овцы), ведет к появлению вызванных потенциалов в обширных территориях мозговой коры (рис. 27).
Рис. 27. Территория появления вызванных потенциалов в коре головного мозга свиньи при раздражении пятачка (по Эдриану)
Таким образом, факты показали, что функциональную карту мозговой коры можно составить с помощью не только метода непосредственного раздражения ее участков, но и метода непрямого — естественного или электрического (в частности, метода вызванных потенциалов) и что этот путь исследования также подтверждает принцип функциональной организации коры, о котором мы уже упоминали.
Метод вызванных потенциалов помогает сделать еще один существенный шаг в прослеживании более сложных форм работы основных систем головного мозга.
Как показал ряд наблюдений (Линдсли, 1961; Гарсиа-Аустт, 1963; И.А.Пеймер, 1958, 1966; Д.А.Фарбер, 1969), сложные воздействия (рассматривание сложных фигур и т.п.) вызывают в мозговой коре существенно иные электрофизиологические процессы (вызванные ответы) по сравнению с относительно более простыми. При этом меняются как латентные периоды вызванных потенциалов, так и их конфигурация.
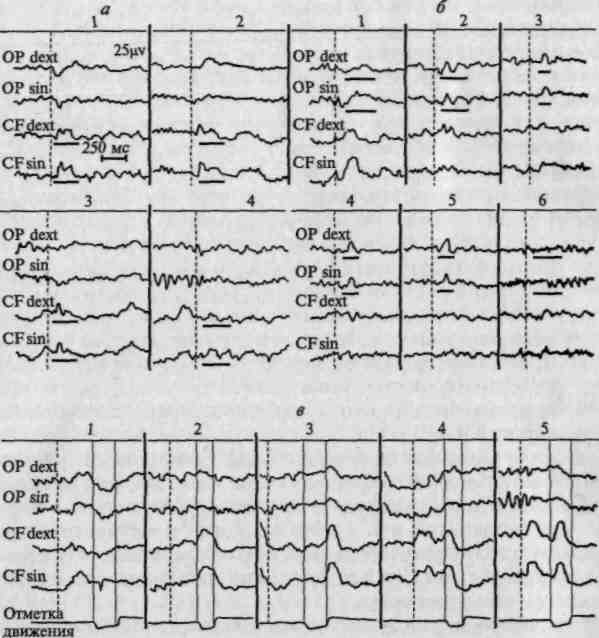
На рисунке 28 мы приводим иллюстрацию этого положения, заимствованную из работы Э. Г. Симерницкой, проведенной в нашей лаборатории. Как показали эти опыты, у нормальных взрослых испытуемых зрительные стимулы порождают вызванные потенциалы только в затылочно-теменных отделах коры, причем эти потенциалы появляются с латентным периодом 80 — 100 мс; в отличие от этого задача дифференцировать предъявляемые зрительные стимулы по длительности приводит к появлению вызванных потенциалов не только в затылочно-теменных, но и в центрально-лобных отделах коры, причем эти потенциалы имеют значительно больший латентный период (от 120 до 260 мс) (Э. Г. Симерницкая, 1970).
Рис. 28. Изменение вызванных потенциалов (ВП) в зависимости от изменения задачи. Динамика вызванных потенциалов при изменении сигнального значения раздражителя у здоровой испытуемой. Ожидание болевого раздражителя (а, 2, 3) вызывает локальные изменения ВП в передних отделах полушарий, проявляющиеся в укорочении временных параметров ВП в передних отделах полушарий по сравнению с «фоновыми» показателями (а, 1). При отмене инструкции (а, 4) исчезает возникшее изменение ВП. Различение зрительных стимулов по длительности приводит к отчетливым изменениям ВП в затылочно-теменных областях коры (б, 2 —5) в виде резкого укорочения цикла ВП. Отмена инструкции (а, 6) снимает эффект активации. Инструкция, требующая по возможности быстрее реагировать двигательной реакцией в ответ на появление зрительного стимула, вызывает четкие изменения ВП в двигательных областях коры, показывая, что ВП могут быть в разных отношениях к моменту ответного действия, опережая его (в, 1), совпадая с ним (в, 2) и даже обнаруживаясь после его завершения (в, 3). По мере автоматизации двигательных навыков в центрально-лобных отведениях начинают регистрироваться две высокоамплитудные волны, одна из которых предшествует двигательному ответу, а другая — сопровождает его (в, 4, 5) (по Э. Г. Симерницкой)
Все это показывает, что метод вызванных потенциалов позволяет получать факты, убедительно говорящие о том, что более сложные формы активной психической деятельности вызывают более обширные процессы в коре головного мозга, вовлекая в совместную работу более сложные системы корковых зон.
Далее мы увидим, какое большое значение имеет это положение для понимания основных форм работы человеческого мозга как органа психической деятельности.
Опыты с анализом функций отдельных нейронов
Метод вызванных потенциалов позволил сделать еще один существенный шаг по пути познания наиболее глубоких, интимных механизмов работы головного мозга, полное значение которого будет оценено, возможно, только следующим поколением исследователей.
Развитие экспериментальной нейрофизиологии за последние десятилетия дало возможность перейти от объективного анализа функций различных систем мозговой коры к изучению функций ее составляющих элементов — отдельных нейронов.
Вживление под контролем специальных стереотактических приборов тончайших электродов в кору головного мозга или подкорковые образования и последующее отведение токов действия от отдельных нейронов при предъявлении животному различных раздражителей позволило Юнгу (1958, 1961), Хьюбелу и Визелу (1962, 1963), а также другим авторам обнаружить, что нейроны реагируют лишь на строго избирательные раздражители. Так, в зрительной коре головного мозга были обнаружены нейроны, которые реагируют пачкой импульсов только на включение или только на выключение света. Были обнаружены также нейроны, которые реагируют как на световые, так и на звуковые или кинестетические раздражители и функция которых носит, таким образом, сложный, мультимодальный характер. Наконец, были обнаружены нейроны, которые не реагируют ни на один сенсорный раздражитель и имеют, следовательно, совсем другие функции.
Аналогичные факты были получены и при исследовании нейронов височной (слуховой) коры. Так, Кацуки (1962), Эванс и др. (1965) обнаружили в слуховой коре нейроны, которые реагируют только на высокие или только на низкие тона, причем распределение этих нейронов имело топически строго упорядоченный характер. Такие же данные были получены Маунткастлом (1957, 1959, 1966) при исследовании соматосенсорной коры.
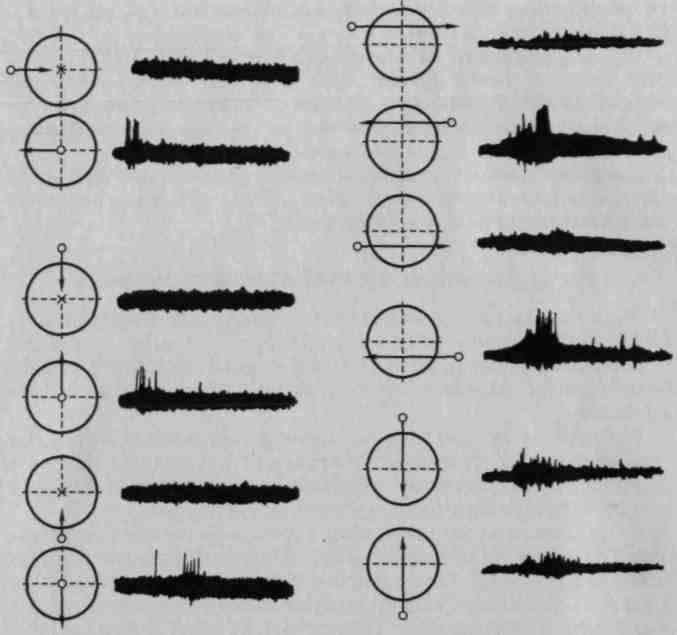
Подробное изучение данных нейронов дало возможность убедиться в их высочайшей специализации. Исследования обнаружили в коре (и в соответствующих подкорковых образованиях низших позвоночных) нейроны, которые реагируют только на прямые или только на плавные закругленные линии, только на острые углы или только на округлые очертания, только на движения точки от центра к периферии или только от периферии к центру и т.д. (рис. 29).
Рис. 29. Высокоспециализированные нейроны зрительных образований золотой рыбки, реагирующие на различные по направлению движения точки (по Кросби-Димону)
Исследование нейронов, входящих в состав вторичных зон коры, выявило существенно иную картину. С одной стороны, в этих областях были обнаружены нейроны, специфически реагирующие на более комплексные признаки, например, на наклон и толщину линии, на конфигурацию линий (Хьюбел и Визел, 1965). С другой стороны, во вторичных зонах коры было обнаружено значительное число нейронов, реагирующих на различные по модальности сигналы, а также нейроны, которые не реагируют ни
на какие сенсорные стимулы и, по-видимому, имеют совсем иную функцию. В самое последнее время во вторичных зонах зрительной коры кошки были выделены нейроны, реагирующие на определенное количество стимулов, причем некоторые из этих нейронов реагируют на это число как при зрительном, так и при слуховом предъявлении (Томпсон, Майерс и др., 1970). Таким образом, уже первые данные позволяют предполагать, что вторичные зоны коры включают в себя нейроны, имеющие значительно более сложные функции по сравнению с нейронами первичных зон.
Многообразие функций отдельных нейронов мозговой коры не ограничивается, однако, только что приведенными характеристиками. Юнг (1958, 1961) первым выделил в коре нейроны, которые не реагировали ни на какие сенсорные раздражители, и предположил, что они имеют какие-то иные функции.
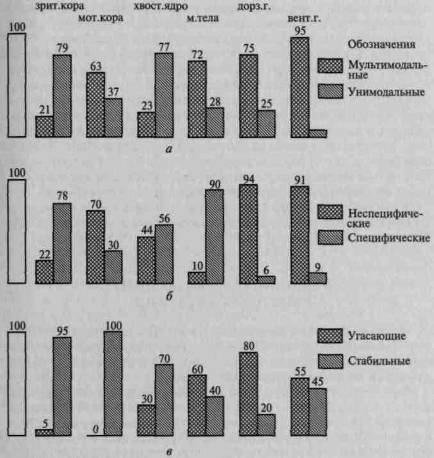
Дальнейшие исследования, проведенные Джаспером (1963), Е.Н.Соколовым (1959), О.С.Виноградовой (1968, 1970), показали, что многие из этих нейронов, не реагируя на специальные сенсорные раздражители, активно реагируют на смену раздражителей или изменение каких-нибудь их свойств, снижая свою активность по мере привыкания и снова активизируясь при появлении изменений (рис. 30). Это заставило исследователей предположить, что они осуществляют функцию сличения новых раздражителей со следами от старых. Оказалось, что эти нейроны, получившие название нейроны внимания (attention units), распределены по мозговым образованиям очень неравномерно: если в первичных зонах зрительной коры их относительно мало, то в образованиях лимбической области, гиппокампе, хвостатом ядре, а у человека, по-видимому, и в наиболее сложных корковых формациях лобной области коры нейроны такого типа встречаются несравненно чаще.
Рис. 30. Распределение нейронов различного типа в разных структурах мозга, % (по О.С.Виноградовой): а — мультимодальные и унимодальные нейроны; б — неспецифические и специфические нейроны; в — угасающие и стабильные нейроны
Все это показывает, что в реализации активных форм психической деятельности, требующих не только получения информации, но и сличения этой информации с предшествующим опытом, принимают участие различные, в том числе далеко отстоящие друг от друга, зоны коры головного мозга и что психические процессы осуществляются сложными системами совместно работающих зон мозговой коры и нижележащих нервных образований.
Значение этого факта для понимания основных принципов функциональной организации мозга как органа психической жизни будет показано далее.
3 ФИЗИОЛОГИЧЕСКИЕ ДАННЫЕ: МЕТОД РАЗРУШЕНИЯ
Мы коротко остановились на двух источниках наших знаний о функциональной организации мозга: сравнительно-анатомических исследованиях и методах раздражения различных участков мозга непосредственным и косвенным путем.
Нам осталось ознакомиться с основными данными, полученными третьим путем, — методом выключения (или разрушения), который заключается в том, что исследователи разрушают определенные области мозга животного и прослеживают изменения в его поведении. Этому методу (или, более точно, наблюдениям над больными, у которых ранение, кровоизлияние или опухоль разрушили определенные участки мозга) суждено было сыграть основную роль в становлении новой науки о функциях мозга — нейропсихологии.
Еще на первых этапах физиологического анализа функции мозга было показано, что разрушение различных его участков приводит к далеко не одинаковым результатам.
Так, уже в середине XIX века было твердо установлено, что разрушение той области мозга собаки, которая содержит гигантские пирамидные клетки и, следовательно, соответствует передней центральной извилине человеческого мозга, вызывает паралич противоположных конечностей, в то время как разрушение других участков мозга (этого же полушария) не приводит к такому эффекту. Аналогичные данные были получены в опытах с высшими млекопитающими — обезьянами. Правда, эти опыты с первых же шагов выявили определенные трудности, источники которых стали понятными лишь значительно позже.
Еще в первой половине XIX века Флуранс (1842) показал, что точное ограничение двигательных функций определенными зонами мозговой коры весьма относительно: перешивая накрест нервы, идущие у петуха от определенных зон мозга к крыльям, он не наблюдал никаких изменений в двигательных функциях последних. Через некоторое время Гольц (1876—1881), разрушивший двигательные участки мозговой коры собаки, пришел к выводу, что моторные функции не ограничены у нее строго определенными участками мозга, что нарушенные функции конечностей быстро восстанавливаются после таких операций и что разрушение различных участков мозга приводит скорее к общему снижению активности поведения животного, чем к выпадению специальных, изолированных функций. Представления об отсутствии четкой функциональной организации мозговой коры у животных, существенно противоречащие всем тем фактам, которые мы приводили ранее, длительное время сохранялись в науке. Выдающийся американский исследователь К. С.Лешли (1929) описал факты, которые заставляли думать, что у крысы функции поведения связаны скорее с массой сохранившегося мозга, чем с локализацией разрушенного в опыте мозгового вещества.
Объяснить это противоречие оказалось возможным много позже, когда больше стало известно о тонком строении мозговой коры различных животных и когда были проведены сравнительные исследования, показавшие, к каким различным результатам приводит разрушение одной и той же мозговой ткани у представителей различных этапов развития животного мира.
Популярное:
|
Последнее изменение этой страницы: 2016-08-24; Просмотров: 521; Нарушение авторского права страницы