|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Отношение количества нейронов к одному нервному волокнуСтр 1 из 6Следующая ⇒
в отдельных образованиях мозга на последовательных ступенях эволюции, %
Мы видим, таким образом, что в процессе эволюции удельный вес коры — по сравнению с нижележащими подкорковыми образованиями — непрерывно возрастает.
Естественно, что большие полушария головного мозга и его кора становятся у человека важнейшим аппаратом регуляции поведения. Существенным является тот факт, что это огромное увеличение объема и массы мозга связано не с ростом наиболее древних, стволовых, отделов мозга, а в первую очередь с развитием больших полушарий и их наиболее существенной части — коры (рис. 3).
Рис. 3. Соотношение древних и новых образований мозга на последовательных этапах эволюции (по Шпатиу)
Было бы неправильным думать, что все области коры человеческого мозга развиваются в процессе эволюции равномерно. Внимательный анализ показал, что развитие больших полушарий связано прежде всего с ростом новых областей коры, которые у низших млекопитающих едва намечены, а у человека составляют основную часть коры (табл. 3); древние области коры — палеокортекс (включающий образования коры, еще не отделенные от подкорковых образований), архикортекс (образования двуслойной древней коры, входящей в систему обонятельного мозга) и межуточная кора (образования, носящие переходный характер между только что упомянутыми), напротив, у человека составляют лишь незначительную часть коры, в то время как у низших млекопитающих они доминируют.
Таблица 3 Относительное изменение различных формаций коры головного мозга на последовательных этапах филогенеза, % (по И. Н. Филимонову, 1949)
Рис. 4. Изменение в соотношении первичных, вторичных и третичных зон коры на последовательных этапах эволюции: а — мозг ежа; б — мозг крысы; в — мозг собаки; г — мозг низшей обезьяны; д — мозг высшей обезьяны; е — мозг человека. Крупными точками обозначены первичные (центральные) поля ядерных зон. Точками средней величины — вторичные (периферические) поля ядерных зон. Мелкими точками — третичные поля (зоны перекрытия) (по Г.И.Полякову)
С переходом от высших млекопитающих (обезьян) к человеку эволюция мозга связана преимущественно с увеличением площади наиболее сложных (третичных) зон коры; площадь более элементарных отделов коры (первичных и вторичных) практически не увеличивается (а иногда даже становится меньше) (рис. 4).
В таблице 4 мы приводим данные отечественных исследователей, объединенные Московским институтом мозга.
Эти данные убедительно показывают принципиальные изменения в соотношении отдельных зон мозговой коры при переходе от низших обезьян к высшим, а затем к человеку. Они позволяют видеть, что относительный размер более просто организованной лимбической области снижается с переходом к человеку; относительный размер прецентральной (двигательной) коры остается без изменений; размер первичной (проекционной) затылочной коры у человека даже уменьшается по сравнению с таковой у обезьяны, в жизни которой зрительное восприятие играет особенно большую роль. Напротив, размеры височной области у человека значительно увеличиваются, а размеры третичных полей коры — нижнетеменной и лобной областей — возрастают в несколько раз.
Таблица 4 Относительное изменение поверхностей отдельных зон коры к величине всей коры у высших млекопитающих и человека, % {по С.М.Блинкову, 1955; И.А.Станкевич, 1955; И.Н.Филимонову, 1949; Е.П.Кононовой, 1962; и др.)
* Данные о размерах лобных долей вычислены применительно к макаке.
Мы видим, таким образом, какое огромное место в коре мозга человека отводится аппаратам, связанным с приемом, переработкой (кодированием) и синтезом информации, получаемой от различных анализаторов, и аппаратам, принимающим участие в выработке и сохранении сложнейших программ поведения и контроля психической деятельности.
На детальном анализе роли этих аппаратов в структуре психических процессов человека мы остановимся далее.
Было бы, однако, глубоко неправильным думать, что если у человека кора головного мозга приобретает ведущую роль, то все нервные образования, которые на более низких этапах эволюции были единственными аппаратами, обеспечивающими организацию поведения, теперь совершенно отстраняются от работы.
Важнейший принцип работы мозга заключается в том, что прежние нервные аппараты сохраняются в нем, но сохраняются, если пользоваться выражением Гегеля, в снятом виде, иначе говоря, сохраняются, уступая ведущее место новым образованиям и приобретая иную роль. Они все больше и больше становятся аппаратами, обеспечивающими фон поведения, принимающими активное участие в регуляции состояний организма, передавая как функции получения, переработки и хранения информации, так и функции
создания новых программ поведения и регуляции и контроля сознательной деятельности высшим аппаратам коры головного мозга (рис. 5).
Рис. 5. Соотношение различных функциональных уровней нервной системы: м — кора мозжечка; зб — зрительный бугор; дек — комплекс филогенетически более старых формаций коры; и — филогенетически более старая часть подкорковых узлов больших полушарий; с — филогенетически более новая часть подкорковых узлов больших полушарий; кп — наиболее поздно появляющиеся в эволюции позвоночных формации полноразвитой новой коры больших полушарий; пи — пирамидный путь для проведения к рефлекторным центрам координационного механизма формируемых в коре импульсов произвольных движений; эпи — экстрапирамидный путь для проведения влияния коры больших полушарий на кору мозжечка. Низшие рефлекторные центры спинного мозга и стволовой части головного мозга представлены черными кругами и треугольниками. Римские цифры соответствуют: I — координационному механизму; II — анализаторно-координационному механизму; III и IV — двум последовательным ступеням прогрессивного усложнения высших (надосевых) мозговых концов систем анализаторов (несколько видоизмененная схема Н.А.Бернштейна)
Забывать это положение и рассматривать аппараты коры головного мозга в отрыве от лежащих ниже образований означало бы допускать грубейшую ошибку. Мы хорошо знаем сейчас, что разные по сложности формы поведения даже у человека могут осуществляться с помощью различных уровней нервной системы.
Каждый физиолог и невролог хорошо знает, что такие простейшие элементы поведения, как сегментарные рефлексы (например, коленный рефлекс, элементарные защитные рефлексы), осуществляются лишь механизмами спинного мозга, и у больного, у которою ранение полностью отделило аппараты спинного мозга от более высоких уровней, эти рефлексы могут сохраняться или даже усиливаться, хотя и не улавливаются сознанием.
Физиологам известно также, что такая сложнейшая врожденная форма поведения, как регуляция обменного равновесия (го-меостаза), обеспечиваемая дыханием, пищеварением и терморегуляцией, осуществляется посредством механизмов, заложенных в верхних отделах ствола (продолговатом мозге, гипоталамусе); при нарушении их соответствующие процессы расстраиваются; грубые поражения этих механизмов могут привести к нарушению «витальных функций» и смерти.
Кроме того, физиологи и неврологи знают, что еще более сложные формы поведения, предполагающие обеспечение тонуса, синергий и координацию, тесно связаны с работой межуточного мозга и подкорковых двигательных узлов (таламо-спинальной системы); поражение их, не вызывая нарушения сложных познавательных процессов, приводит к грубому нарушению «фонового» поведения. Особый интерес в связи с этой проблемой представляют результаты наблюдений над больными, страдающими паркинсонизмом, накопившиеся за последние три десятилетия в результате широкого изучения эпидемического энцефалита и распространения стереотактических операций.
Наконец, хорошо известно, что наиболее сложные формы деятельности не могут быть обеспечены без участия коры головного мозга, являющейся органом высших форм поведения животных и сознательного поведения человека.
Таким образом, ясно, что сложные рефлекторные процессы и сложные формы поведения могут осуществляться разными уровнями нервной системы, каждый из которых вносит в функциональную организацию поведения свой вклад.
Последние десятилетия позволили во многом уточнить только что обозначенное положение. Было показано, что низшие уровни нервного аппарата участвуют в организации работы коры больших полушарий, регулируя и обеспечивая ее тонус.
Роль нижних отделов ствола и образований межуточного мозга в обеспечении и регуляции тонуса коры была показана сравнительно недавно благодаря классическим работам Мэгуна и Моруцци (1949 и др.), посвященным так называемой восходящей активирующей ретикулярной формации.
Забывать об участии низших уровней мозгового аппарата в наиболее сложных формах поведения и игнорировать тот факт, что они обеспечивают нужное состояние коры и выступают как регулятор общего фона психической деятельности, значило бы сейчас допускать серьезную ошибку. Далее (ч. первая, гл. III) мы остановимся на этом вопросе подробнее.
Аппараты стволового уровня не работают в полной изоляции от коры головного мозга и сами испытывают ее регулирующее влияние.
Работы Мак-Кэллока и др. (1946), Френча и др. (1955), Линдсли (1955, 1956, 1961), Жувэ и др. (1956, 1961), Эрнандес-Пеона (1966, 1969) и большое число исследований, на которых мы еще остановимся далее, показали огромную роль нисходящей активирующей ретикулярной формации, направляющей импульсы от коры головного мозга к нижележащим образованиям и приводящей аппараты регуляции тонуса в соответствие с информацией, получаемой субъектом, и с задачами, которые он ставит перед собой.
Данные, полученные в современных анатомических и физиологических исследованиях, позволяют сформулировать принцип вертикального строения функциональных систем мозга, иначе говоря, принцип, согласно которому каждая форма поведения обеспечивается совместной работой разных уровней нервного аппарата, связанных друг с другом как восходящими (летальными), так и нисходящими (фугальными) связями, превращающими мозг в саморегулирующуюся систему.
Этот прочно вошедший в науку принцип утверждает, что кора головного мозга, находящаяся в постоянном взаимодействии с нижележащими образованиями, не является единственным мозговым субстратом психических процессов.
Этот принцип делает понятным и те факты, которые ставили в тупик многих исследователей прошлого. Было показано, что разобщение отдельных зон коры путем круговой изоляции может не влечь за собой существенных изменений в поведении животных, в то время как подрезка коры, изолирующая ее от нижележащих образований, неизбежно приводит к значительным нарушениям ее регулирующих функций (Чоу, 1954; Сперри, 1959; Прибрам, Блейрт, Спинелли, 1966). Все это означает, что отдельные участки коры головного мозга соединяются между собой не только с помощью горизонтальных (транскортикальных) связей, но и через нижележащие образования, иначе говоря, посредством системы вертикальных связей.
Имея в виду сформулированные выше положения об эволюции нервных аппаратов, основных уровнях нервной системы и их взаимодействии, мы переходим сейчас к рассмотрению данных, которыми располагает сравнительная анатомия коры головного мозга.
Структурная и функциональная организация коры головного мозга
Наблюдения, показавшие, что мозг в целом и его кора в частности обладают неоднородным строением, относятся еще к началу прошлого века.
Ф. Галль, известный анатом, вошедший в историю науки как основатель фантастической «френологии» (концепции о функциональной организации мозга, исходящей из представлений о локализации сложных психических «способностей» в его ограниченных участках), впервые отличил серое вещество, составляющее мозговую кору и подкорковые серые образования, от белого вещества, состоящего из проводящих волокон, связывающих отдельные участки коры и соединяющие кору большого мозга с периферией. Однако это открытие, сделавшее Галля подлинным основателем морфологии мозга, долго оставалось без адекватной оценки, и настоящее раскрытие функций коры головного мозга, ее проводящих путей и серого вещества, заложенного в глубине больших полушарий, было сделано лишь спустя несколько поколений.
Значительный шаг вперед был сделан в 1863 г. киевским анатомом В. А. Бецом, занимавшимся микроскопическим изучением клеточного состава мозговой коры. Ему принадлежит открытие, которому было суждено стать началом целой эпохи блестящих исследований.
Описывая строение различных участков мозговой коры, он обнаружил, что их морфологическая структура в высокой степени неоднородна: если кора передней центральной извилины включает в свой состав большие, имеющие форму пирамиды нервные клетки (они получили в дальнейшем название гигантских пирамидных клеток Беца), то прилегающая к ней кора задней центральной извилины имеет совсем иное, мелкозернистое строение и совсем лишена пирамидных клеток (рис. 6).
Рис. 6. Два вида строения коры: а — передняя (моторная) кора; 6 — задняя (сенсорная) кора (по Бродману)
Позднее было установлено, что различие этих двух областей коры не только морфологическое, но и функциональное. Гигантские пирамидные клетки Беца (составляющие пятый слой коры) оказались источниками двигательных импульсов, идущих от коры к периферической мускулатуре, а передняя центральная извилина, в которой они были сосредоточены, — моторной областью коры головного мозга. Поля мозговой коры, имеющие мелкозернистое строение и отличающиеся развитым четвертым слоем нервных клеток (к их числу относятся и образования задней центральной извилины), оказались аппаратами, к которым подходят чувствительные волокна, начинающиеся в периферических органах чувств (рецепторах), а соответствующие зоны коры — первичными чувствительными образованиями коры большого мозга.
С выделением двигательных и сенсорных областей (или первичных двигательных и сенсорных центров) был сделан первый шаг к созданию функциональной карты коры головного мозга, и кажущаяся однородной масса серого вещества, покрывающая тонким слоем большие полушария, начала приобретать дифференцированный характер.
Дальнейшие сравнительно-анатомические наблюдения подтвердили плодотворность наметившегося подхода. Оказалось, что внимательное изучение «первичных» областей мозговой коры позволяет делать точные выводы о некоторых особенностях поведения животного. Следующие примеры хорошо иллюстрируют это положение.
Рис. 7. Строение двигательной коры: а — австралийского медвежонка; б — летучей собаки (по Бродману)
На рисунке 7 мы приводим срезы из двигательных областей мозговой коры австралийского медвежонка (рис. 7, а) и летучей собаки (рис. 7, б). Легко увидеть, что на первом срезе присутствует сравнительно немного гигантских пирамидных клеток, в то время как на втором срезе число их значительно больше, а величина гораздо меньше. Не указывает ли нам этот факт на относительно сильные и грубые движения первого животного и более тонкие и многообразные движения второго?
К аналогичным заключениям позволяет прийти и сравнительно-анатомический анализ строения сенсорных отделов коры.
Рис. 8. Протяженность обонятельной коры: а, б — большой мозг ежа с боковой и медиальной сторон; в, г — большой мозг человека. Новая кора оставлена незаштрихованной. У ежа она занимает 1/4, а у человека 11/12 всей поверхности мозга (по Экономо)
Рисунок 8 показывает то место, которое занимают образования обонятельных полей в коре головного мозга ежа (рис. 8, а, б) и человека (рис. 8, в, г). Он указывает на ведущую роль обонятельного анализатора у низших млекопитающих и незначительное место этого анализатора у человека.
Подобный вывод можно сделать и при сравнении зрительных полей животных и человека (рис. 9): значительное развитие мелкоклеточных образований зрительной коры обезьяны, составляющих у нее до 40 % площади коры (рис. 9, б), по сравнению с такими же образованиями зрительной коры крота (рис. 9, а) объясняется тем, что ведущее место в поведении первого животного занимает зрение, в то время как в поведении второго животного, ориентирующегося в окружающем мире с помощью обоняния, оно занимает лишь сравнительно небольшое место.
Рис. 9. Протяженность зрительной коры: а — крота; б — обезьяны; в — человека (пунктиром обозначены мелкоклеточные образования первичной зрительной области коры) (по Бродману)
Сравнительно-анатомическое изучение коры головного мозга, начавшееся с выделения основных, первичных, или проекционных, зон мозговой коры, существенно продвинулось за последние десятилетия.
Решающие успехи были сделаны еще в начале этого столетия, когда работы Кэмпбелла (1905) и Бродмана (1909), Рамон-и-Кахала (1909 — 1911) и Болтона (1933) позволили составить цитоархитектонические карты мозговой коры; эти карты, существенно уточненные Ц. и О. Фогтами (1919 — 1920) и работами Московского института мозга, позволили приблизиться к описанию основных принципов строения мозговой коры животных и человека и внесли неоценимый вклад в наши знания о мозге как органе психики.
Как показали эти исследования, новая кора головного мозга состоит из шести слоев клеток (рис. 10). Только нижние из них являются аппаратами, непосредственно связывающими мозговую кору с периферией: органами чувств (IV — афферентный слой) и мышцами (V — эфферентный слой). В IV слое коры приходят волокна, несущие импульсы, возникающие в периферических ре-
цепторах; именно этот слой мелкозернистых клеток особенно мощно развит в только что упомянутых первичных чувствительных зонах коры. V слой включает в себя гигантские пирамидные клетки, генерирующие импульсы к мышцам тела и дающие начало длинному двигательному пути, состоящему из нервных волокон; абсолютное преобладание этого слоя имеет место в передней центральной извилине, или двигательной зоне, коры головного мозга.
Рис. 10. Типы архитектонического строения коры больших полушарий человека: а — тип двигательной области; б — фронтальный тип; в — париетальный тип; г — полярный тип; д — гранулярная, зернистая кора; I—V — слои коры (по Экономо)
На рисунке 11 мы приводим схему, позволяющую проследить ход волокон от периферических органов чувств в соответствующие «проекционные» отделы коры головного мозга. Она показывает, что волокна, начинающиеся от чувствительных аппаратов кожи и мышц, прерываясь в подкорковых образованиях, приходят к коре задней центральной извилины (общечувствительная зона), а волокна, идущие от сетчатки глаза и от внутреннего уха, также переключаясь в подкорковых аппаратах, заканчиваются соответственно в затылочных и в первичных височных отделах коры.
Рис. 11. Афферентные пути и чувствительные зоны коры. Жирными линиями показаны системы анализаторов с их переключениями в подкорковых отделах: / — зрительный анализатор; 2 — слуховой анализатор; 3 — кожно-кинестети- ческий анализатор. Т — височная область; О — затылочная область; Pip — поле 39; Pia — поле 40; Pstc — постцентральная область; ТРО — височно- теменно-затылочная область; ТИ — зрительный бугор; Cgm — внутреннее коленчатое тело; Cgl — наружное коленчатое тело (по Г. И. Полякову)
Таким образом, в коре головного мозга человека выделяются проекционная общечувствительная (теменная), зрительная (затылочная) и слуховая (височная) области.
Аналогичным образом мы можем проследить волокна, которые, начинаясь в передней центральной извилине и подходя к передним рогам спинного мозга, несут двигательные импульсы к мышцам. Эти волокна составляют двигательный, или пирамидный, путь головного мозга.
Как показали морфологические исследования, над каждой «первичной» зоной коры (с преобладающим развитием IV — афферентного или V — эфферентного слоев клеток) надстраивается система «вторичных» зон, в которых преобладающее место занимают более сложные по своему строению II и III слои. Эти слои состоят из клеток с короткими аксонами, большая часть которых или не имеет прямой связи с периферией, или получает свои импульсы из лежащих в глубине мозга подкорковых образований, осуществляющих первичную переработку приходящих с периферии импульсов. Строение этих слоев позволяет относить их уже не к простейшему — «проекционному», а к гораздо более сложному — «ассоциативному», или «интегрирующему», аппарату коры головного мозга.
Существенным для понимания функции этих слоев коры головного мозга является тот факт, что в процессе эволюции видов удельный вес их непрерывно увеличивается (рис. 12); это показывает, что процесс усложнения психической деятельности, переход от относительно простых, врожденных форм поведения животного к более сложным формам кодирования поступающей информации у человека, предполагающим сознательный характер программирования деятельности, связаны с развитием этих высших слоев мозговой коры.
Рис. 12. Сравнительная толщина верхних слоев коры в филогенезе: а — срезы; б — схема (по данным Московского института мозга)
Другой не менее важной функциональной характеристикой строения коры мозга животного является отношение между массой клеточных тел и массой клеточного вещества.
Исследования последнего времени показали, что в осуществлении сложных нервных процессов решающую роль играет не только тело нервной клетки, но и ее многочисленные отростки и, наконец, окружающие нейроны глиальные клетки (Хиден, 1962, 1964; Ройтбак, 1965; и др.).
Увеличение «глиального индекса» на каждой новой ступени эволюции указывает на повышение управляемости функций отдельных мозговых зон, однако лишь будущие сравнительно-анатомические исследования могут выявить его недлинный функциональный смысл.
Характерным поэтому является тот факт, что с эволюцией животного величина отношения глиальной ткани коры к массе ее нервных клеток все более возрастает и у человека оказывается во много раз большей, чем у млекопитающих, стоящих на более низких ступенях эволюции (табл. 5).
Таблица 5 А. Отношение массы серого вещества коры (нейронов) к массе глнальных клеток («глиальный индекс») на разных ступенях эволюции (по Фрид, 1954)
Б. Отношение массы серого вещества коры (нейронов) к массе глнальных клеток («глиальный индекс») на последовательных ступенях эволюции в пределах двигательной зоны коры (по Банану, 1951)
Аналогичная тенденция легко прослеживается в процессе созревания коры мозга человека. У плода 6 месяцев верхние слои коры едва намечены, у младенца — развиты относительно слабо, у нормального взрослого — занимают значительное место (рис. 13). В случаях врожденного слабоумия эти слои клеток недоразвиты, а у больных с органической деменцией и атрофией коры — резко сужены.
Рис. 13. Сравнительная толщина верхних слоев коры в онтогенезе (по Е. П. Кононовой и др.)
Все это указывает на то, что верхние, «ассоциативные», слои мозговой коры играют важную роль в осуществлении наиболее сложных форм психической деятельности, становление которых происходит на поздних ступенях филогенеза и на поздних этапах развития человека.
Не менее важным анатомическим фактом, позволяющим нам понять основные принципы строения мозговой коры, является неравно-
мерность распределения отдельных слоев коры в топографически различных участках коры головного мозга.
Факты показывают, что над каждой первичной областью мозговой коры, в которой преобладают низшие (афферентные или эфферентные) слои коры, надстраиваются вторичные области, в которых преобладают верхние (проекционно-ассоциационные) слои, играющие, как мы увидим далее, существенную роль в функциональной организации работы отдельных анализаторов.
Иерархическое строение мозговой коры легко можно видеть, рассмотрев топографическую карту мозга, изображенную на рисунке 14. Мы видим, что над первичными (проекционными) отделами общечувствительной коры (задняя центральная извилина) у человека надстраивается вторичная чувствительная кора, в которой преобладают верхние (проекционно-ассоциационные) слои; над первичной зрительной корой, расположенной в полюсе затылочной области, надстраивается вторичная зрительная кора, где также преобладают верхние (проекционно-ассоциационные) слои; над первичной слуховой корой, расположенной в верхних отделах височной области, надстраиваются ее вторичные отделы с тем же строением; наконец, над первичной двигательной корой, занимающей переднюю центральную извилину, — ее вторичные отделы, расположенные в премоторной области.
Рис. 14. Топографическая карта мозговой коры: а — наружная поверхность; б — внутренняя поверхность. Ядерные зоны мозговой коры обозначены кружками (зрительная зона), квадратами (слуховая зона), ромбами (общечувствительная зона), треугольниками (двигательная зона); центральные поля выделены более крупными знаками. Зоны перекрытия анализаторов в задних отделах полушария (теменно-височно-затылочные и нижнетеменные отделы) обозначены смешанными знаками; в передних отделах полушария (лобная область) — видоизмененными треугольниками; лимбическая и инсулярная области, а также филогенетически старые зоны коры — прерывистой штриховкой (по Г.И.Полякову) 2 Лурия
Как видно из той же карты, в коре головного мозга человека можно выделить участки, которые лежат на границах между корковыми представительствами отдельных чувствительных зон мозговой коры и которые получили название третичных зон коры (или зон перекрытия коркового представительства отдельных анализаторов). Эти области коры целиком состоят из верхних (ассоциационных) слоев клеток и не имеют прямой связи с периферией. Есть все основания предполагать, что третичные зоны коры обеспечивают совместную работу корковых звеньев отдельных анализаторов, наиболее сложные интегральные функции коры головного мозга.
Как показали детальные анатомические исследования, в коре головного мозга можно выделить две группы третичных областей. Первая из них — задняя — расположена на стыке зрительной (затылочной), общечувствительной (теменной) и слуховой (височной) областей; ее с полным основанием можно обозначить как зону перекрытия корковых отделов экстероцептивных анализаторов. Вторая — передняя — расположена спереди от двигательной зоны коры и надстраивается над двигательными отделами коры головного мозга. Она связана со всеми остальными отделами коры и, как мы увидим далее, играет существенную роль в построении наиболее сложных программ поведения человека.
Внимательное изучение хода волокон от периферических органов чувств к коре головного мозга в полной мере подтверждает принцип иерархического строения основных отделов мозговой коры. Рисунки 14 и 15 убедительно показывают, что функциональные системы головного мозга имеют одинаковое (иерархическое) строение и что анатомические данные позволяют выделить в коре головного мозга первичные, вторичные и третичные зоны.
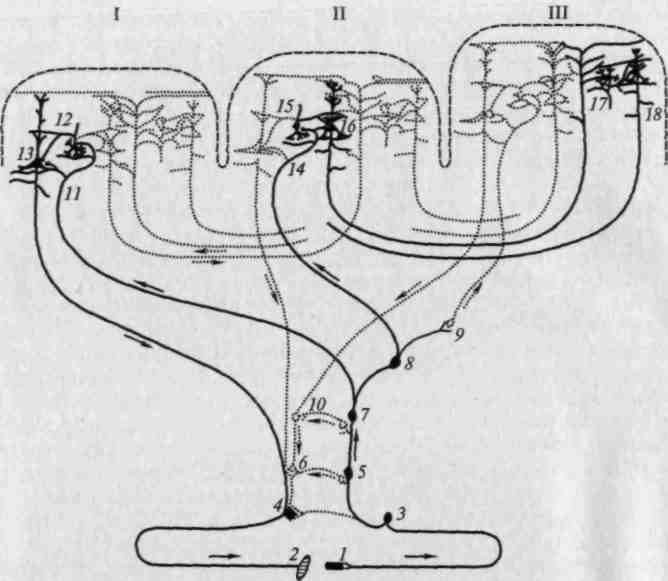
Рис. 15. Системы связей первичных, вторичных и третичных зон мозговой коры: I — первичные (центральные) поля; II — вторичные (периферические) поля; III — третичные поля (зоны перекрытия анализаторов). Жирными линиями выделены: I — система проекционных (корково-подкорковых) связей коры; 11 — система проекционно-ассоциационных связей коры; III — система ассоциативных связей коры. 1 — рецептор; 2 — эффектор; 3 — нейрон чувствительного узла; 4 — двигательный нейрон; 5, 6 — переключательные нейроны спинного мозга и ствола; 7—10 — переключательные нейроны подкорковых образований; 11, 14 — афферентные волокна из подкорки; 13 — пирамида V слоя; 16 — пирамида подслоя III3; 18 — пирамиды подслоев III2 и Ш1; 12, 15, 17 — звездчатые клетки коры (по Г.И.Полякову)
Было бы неверным думать, что три описанных выше типа зон мозговой коры отмечаются на всех ступенях эволюции позвоночных. Факты говорят об обратном и указывают на то, что описанное нами иерархическое строение коры головного мозга является продуктом длительного исторического развития.
Как показывают сравнительно-анатомические данные, схематически представленные ранее на рисунке 4, в коре головного мозга ежа и крысы дифференциация первичных и вторичных зон едва намечается, а третичные зоны коры совсем отсутствуют; близкое строение имеет кора головного мозга собаки, лишь у обезьяны вторичные и третичные зоны мозговой коры отмечаются достаточно ясно. У человека иерархическое строение коры головного мозга выступает с полной отчетливостью; схема на рисунке 14 показывает, что первичные участки мозговой коры занимают у него совсем небольшое место, будучи оттесненными хорошо развитыми вторичными участками, и что третичные зоны мозговой теменно-височно-затылочной и лобной коры становятся здесь наиболее развитыми системами и занимают подавляющую часть коры больших полушарий.
В самое последнее время были получены очень важные данные, помогающие заполнить существенный пробел в наших знаниях о предыстории человеческого мозга.
Мы только что привели данные, касающиеся основных тенденций прогрессивного развития вторичных и третичных зон коры на последовательных этапах эволюции животного мира. Один вопрос остался, однако, неосвещенным: можно ли проследить такую же тенденцию и на последовательных этапах антропогенеза? Знаменуется ли переход от приматов к гоминидам и последующий переход от древних форм предчеловека к более поздним и совершенным формам соответствующим возрастанием удельного веса вторичных и третичных зон коры, отражающим усложнение условий жизни и переход от животной эволюции к социальной истории человека?
До сих пор мы знали лишь то, что объем головного мозга от высших обезьян к человеку постепенно растет и что с переходом к человеческой истории этот рост останавливается. По вычислениям Бонина (1934), средняя емкость черепа кроманьонца равнялась в среднем 1570 см3, приблизительно такая же емкость черепа характерна и для представителей неолита (1525 — 1533 см3), железного века (1514 см3) и различных периодов истории Древнего Египта (1390-1495 см3).
Означает ли это, что на всем протяжении антропогенеза соотношение отдельных образований мозговой коры оставалось неизменным?
До недавнего времени сколько-нибудь обоснованный ответ на этот вопрос казался невозможным. Только в последние годы были достигнуты первые успехи в решении этой проблемы. Связаны они с использованием оригинального метода получения слепков мозга с внутренней поверхности черепа (эндокраниума) различных предков человека. Наряду с учеными других стран большую работу в этом направлении проделала отечественный антрополог
В. И. Кочеткова (1966 и др.). Создавая слепки мозга с внутренней поверхности черепа ископаемого человека и пользуясь некоторыми опорными пунктами (брегма, отпечатки сильвиевой и передней центральной извилин, а также сосудов, оставляющих свой след на внутренней поверхности мозга), она смогла получить приближенные реконструкции, позволяющие определить соотноше-
ния отдельных частей мозга на последовательных этапах праистории (рис. 16). Дальнейший математический подсчет полученных данных позволил Кочетковой сделать некоторые выводы, имеющие самое непосредственное отношение к ответу на интересующий нас вопрос. Кратко остановимся на них.
Рис. 16. Развитие мозга на последовательных этапах антропогенеза (по слепкам с эндокранов): 1 — питекантроп; 2 — синантроп; 3 — неандерталец (Брокен-Хилл); 4 — то же (Ля Шапелль); 5 — то же (Пржедмост); 6 — современный человек (по Тильней и Дюбуа)
Популярное:
|
Последнее изменение этой страницы: 2016-08-24; Просмотров: 424; Нарушение авторского права страницы