|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
АВТОНОМНАЯ САМОРЕГУЛЯЦИЯ ГМФ-ПУТИ.
Лимитирующим звеном ГМФ-пути является реакция окисления 6-фосфоглюконовой кислоты, катализируемая 6-фосфоглюконат-дегидрогеназой. Vmax этого фермента меньше, чем Vmax любого другого фермента ГМФ-пути. Поэтому он определяет собой Vmax всего ГМФ-пути в целом и является лимитирующим ферментом. Но он не является ключевым, потому что не является регуляторным. Регуляторный фермент ГМФ-пути - это глюкозо-6-фосфат-дегидро-геназа. Его Vmax больше, чем у 6-фосфоглюконатдегидрогеназы, поэтому он не является лимитирующим. Но он ингибируется избытком АТФ и своего продукта НАДН2. Значит, в определенных условиях его Vmax может стать меньше, чем Vmax 6-фосфоглюконатдегидрогеназы. В этих условиях лимитирующая роль переходит к глюкозо-6-фосфатдегидрогеназе. Таким образом, роль ключевого фермента ГМФ-пути выполняют сразу 2 фермента - обе дегидрогеназы, которые разделяют между собой функции ключевого и лимитирующего ферментов. Эти ключевые ферменты являются " пунктами вторичного контроля" ГМФ-пути. Скорость ГМФ-пути определяется либо 6-фосфоглюконат-дегидрогеназой, либо (когда много НАДФН2 и АТФ) - глюкозо-6-фосфат-дегидрогеназой. Реакции ГМФ-пути регулируются генетически на уровне биосинтеза этих ферментов. ГМФ-путь существует не во всех типах клеток. Наиболее интенсивно ГМФ-путь протекает в печени, эритроцитах, надпочечниках, половых железах, жировой ткани и молочной железе. Но даже в этих тканях ГМФ-путем расщепляется не более 25-30% глюкозы. Почему в этих тканях ГМФ-путь имеет такое значение? Потому что клеткам этих тканей нужно много НАДФН2. Например: В жировой ткани с большой скоростью идет синтез жиров. В печени - синтез жиров, холестерина. В коре надпочечников и в половых клетках синтезируются стероидные гормоны. Для этого нужно много НАДФН2. Регуляция на генетическом уровне идет медленно, требуется время для проявления эффекта этого типа регуляции. Автономная регуляция приводит к практически мгновенному эффекту. Зачем нужен ГМФ-путь в эритроцитах? Здесь тоже требуется НАДФН2, но не для синтезов, а для защиты от токсических эффектов кислорода. В эритроцитах многие ферменты имеют в активных центрах SH-группы, которые могут окисляться под действием кислорода. Накопленный при распаде глюкозы в ГМФ-пути НАДФН2 позволяет регенерировать эти SH-группы. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ МЕТАБОЛИЗМА УГЛЕВОДОВ ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ЭНЕРГЕТИЧЕСКОГО МЕТАБОЛИЗМА. Действие гормонов, влияющих на энергетический метаболизм можно увидеть при определении некоторых биохимических показателей. Например, концентрации глюкозы в крови. Гормоны делят на: 1. Повышающие уровень глюкозы в крови; 2. Понижающие уровень глюкозы в крови. Ко второй группе относится только ИНСУЛИН. Также гормоны можно разделить на ГОРМОНЫ ПРЯМОГО ДЕЙСТВИЯ на энергетический метаболизм и ГОРМОНЫ КОСВЕННОГО ДЕЙСТВИЯ. Гормоны прямого действия. ИНСУЛИН Основные механизмы действия инсулина: 1. Инсулин повышает проницаемость плазматических мембран для глюкозы. Этот эффект инсулина является главным лимитирующим звеном метаболизма углеводов в клетках. 2. Инсулин снимает тормозящее действие глюкокортикостероидов на гексокиназу. 3. На генетическом уровне инсулин стимулирует биосинтез ферментов метаболизма углеводов, в том числе ключевых ферментов. 4. Инсулин в клетках жировой ткани ингибирует триглицеридлипазу - ключевой фермент распада жиров. Регуляция секреции инсулина в кровь происходит с участием нейро-рефлекторных механизмов. В стенках кровеносных сосудов есть особые хеморецепторы, чувствительные к глюкозе. Повышение концентрации глюкозы в крови вызывает рефлекторную секркцию инсулина в кровь, глюкоза проникает в клетки и ее концентрация в крови снижается. Остальные гормоны вызывают повышение концентрации глюкозы в крови. ГЛЮКАГОН. Относится к белково-пептидным гормонам. Обладает мембранным типом взаимодействия с клеткой-мишенью. Эффект оказывает через аденилатциклазную систему. 1. Вызывает повышение активности гликоген-фосфорилазы. В результате ускоряется распад гликогена. Так как глюкагон оказывает эффект только в печени то можно сказать, что он " гонит глюкозу из печени". 2. Понижает активность гликоген-синтетазы, замедляя синтез гликогена. 3. Активирует липазу в жировых депо. АДРЕНАЛИН. Имеет рецепторы во многих тканях, а механизмы действия у него такие же, как у глюкагона. 1. Ускоряет распад гликогена. 2. Замедляет синтез гликогена. 3. Ускоряет липолиз. ГЛЮКОКОРТИКОСТЕРОИДЫ (ГКС). Относятся к стероидным гормонам, поэтому обладают внутриклеточным типом взаимодействия с клеткой-мишенью. Проникая в клетку-мишень, они взаимодействуют с клеточным рецептором и обладают следующими эффектами: 1. Ингибируют гексокиназу - таким образом они замедляют утилизацию глюкозы. В результате концентрация глюкозы в крови возрастает. 2. Данные гормоны обеспечивают процесс гликонеогенеза субстратами. 3. На генетическом уровне усиливают биосинтез ферментов катаболизма белков. Гормоны косвенного действия СОМАТОТРОПНЫЙ ГОРМОН. 1. Усиливает выделение глюкагона, поэтому наблюдается ускорение распада гликогена. 2. Вызывает активацию липолиза, поэтому способствует утилизации жира в качестве источника энергии. ЙОДСОДЕРЖАЩИЕ ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ. Трииодтиронин (Т3) Тироксин (Т4) Это гормоны - производные аминокислоты тирозина. Обладают внутриклеточным типом взаимодействия с клетками-мишенями. Рецептор Т3/Т4 находится в ядре клетки. Поэтому эти гормоны усиливают биосинтез белков на уровне транскрипции. Среди этих белков - окислительные ферменты, в частности разнообразные дегидрогеназы. Кроме того, они стимулируют синтез АТФаз, т.е. ферментов, которые разрушают АТФ. Для процессов биоокисления требуются субстраты - продукты окисления углеводов и жиров. Поэтому при увеличении продукции этих гормонов наблюдается усиление распада углеводов и жиров. Гиперфункция щитовидной железы называется Базедова болезнь или тиреотоксикоз. Один из симптомов этого заболевания - понижение массы тела. Для этого заболевания характерно повышение температуры тела. В опытах in vitro наблюдается разобщение митохондриального окисления и окислительного фосфорилирования при высоких дозах этих гормонов.
Организм человека существует как единое целое благодаря системе внутренних связей, которая обеспечивает передачу информации от одной клетки к другой в одной и той же ткани или между разными тканями. Без этой системы невозможно поддерживать гомеостаз. В передаче информации между клетками в многоклеточных живых организмах, принимают участие три системы: ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА (ЦНС), ЭНДОКРИННАЯ СИСТЕМА (ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ) и ИММУННАЯ СИСТЕМА. Способы передачи информации во всех названных системах - химические. Посредниками при передаче информации могут быть СИГНАЛЬНЫЕ молекулы. К таким сигнальным молекулам относятся четыре группы веществ: ЭНДОГЕННЫЕ БИОЛОГИЧЕСКИ АКТИВНЫЕ ВЕЩЕСТВА (медиаторы иммунного ответа, факторы роста и др.), НЕЙРОМЕДИАТОРЫ, АНТИТЕЛА (иммуноглобулины) и ГОРМОНЫ.
ХИМИЯ И ОБМЕН ЛИПИДОВ. ЛИПИДЫ - органические вещества, характерные для живых организмов, нерастворимые в воде, но растворимые в органических растворителях и друг в друге.
Группы липидов отличаются по степени гидрофобности. Фосфолипиды и гликолипиды являются полярными липидами. Холестерин занимает промежуточное положение между полярными и абсолютно гидрофобными липидами. Абсолютно гидрофобными являются триглицериды и эфиры холестерина. Большинство липидов (кроме стеринов и некоторых минорных липидов) содержат высшие жирные кислоты (ВЖК). В состав мембран входят только ФОСФОЛИПИДЫ (ФЛ), ГЛИКОЛИПИДЫ (ГЛ) и ХОЛЕСТЕРИН (ХС). ЖИРНЫЕ КИСЛОТЫ, ХАРАКТЕРНЫЕ ДЛЯ ОРГАНИЗМА ЧЕЛОВЕКА Кислота называется жирной, если число углеродных атомов в ее молекуле больше четырех. Преобладают длинноцепочечные жирные кислоты (число атомов углерода 16 и выше). Количество углеродных атомов и двойных связей обозначается двойным индексом. Например: С18: 1 (9-10). В данном случае 18 – число атомов углерода и 1 – количество двойных связей. В скобках указывается местоположение двойных связей (по номерам углеродных атомов). Студенты должны знать количество углеродных атомов и двойных связей в молекулах следующих жирных кислот: 1. С16: 0 - пальмитиновая, 2. С18: 0 - стеариновая, 3. С18: 1 – олеиновая (9: 10), 4. С18: 2 – линолевая (9-10, 12-13), 5. С18: 3 - линоленовая (9-10, 12-13, 15-16), 6. С20: 4 - арахидоновая (5-6, 8-9, 12-13, 15-16). Жирные кислоты, имеющие двойную связь в положении w3, обладают очень выраженным антиатеросклеротическим действием (линоленовая). Особенно много w3-содержащих жирных кислот находится в рыбьем жире и других морепродуктах. В организме человека такие жирные кислоты не синтезируются. Жирные кислоты, входящие в состав организма человека, имеют общие черты строения: 1. Чётное число атомов углерода. 2. Линейная (неразветвлённая) углеродная цепь. 3. Полиненасыщенные жирные кислоты имеют ТОЛЬКО ИЗОЛИРОВАННЫЕ двойные связи (между соседними двойными связями не меньше двух одинарных). 4. Двойные связи имеют только цис-конфигурацию. По количеству двойных связей жирные кислоты можно разделить на НАСЫЩЕННЫЕ (нет двойных связей), МОНОНЕНАСЫЩЕННЫЕ (есть одна двойная связь) и ПОЛИНЕНАСЫЩЕННЫЕ (две или более двойных связей). Свободные жирные кислоты встречаются в очень небольшом количестве. В основном они находятся в составе других липидов. При этом они связаны с другими компонентами липидов сложноэфирной связью (эстерифицированы).
ФОРМЫ СУЩЕСТВОВАНИЯ ЛИПИДОВ В ОРГАНИЗМЕ ЧЕЛОВЕКА 1. Жировые включения, которых больше всего в адипоцитах. Это форма существования триглицеридов. 2. Биомембраны. Они не содержат триглицеридов, зато в них присутствуют фосфолипиды, гликолипиды и холестерин. 3. Липопротеины. Могут включать в себя липиды всех классов. ЛИПОПРОТЕИНЫ Липопротеины - это сферические частицы, в которых можно выделить гидрофобную сердцевину, состоящую из триглицеридов (ТРГ) и эфиров холестерина (ЭХС) и амфифильную оболочку, в составе которой – фосфолипиды, гликолипиды и белки.
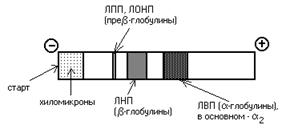
Белки оболочки называются апобелками. Холестерин (ХС) обычно занимает промежуточное положение между оболочкой и сердцевиной. Компоненты частицы связаны слабыми типами связей и находятся в состоянии постоянной диффузии – способны перемещаться друг относительно друга. Основная роль липопротеинов – транспорт липидов, поэтому обнаружить их можно в биологических жидкостях. При изучении липидов плазмы крови оказалось, что их можно разделить на группы, так как они отличаются друг от друга по соотношению компонентов. У разных липопротеинов наблюдается различное соотношение липидов и белка в составе частицы, поэтому различна и плотность. Липопротеины разделяют по плотности методом ультрацентрифугирования, при этом они не осаждаются, а всплывают (флотируют). Мерой всплывания является константа флотации, обозначаемая Sf (сведберг флотации). В соответствии с этим показателем различают следующие группы липопротеинов:
Липопротеины можно разделить и методом электрофореза. При классическом щелочном электрофорезе разные липопротеины ведут себя по-разному. При помещении липопротеинов в электрическое поле хиломикроны остаются на старте. ЛОНП и ЛПП можно обнаружить во фракции преb-глобулинов, ЛНП - во фракции b-глобулинов, а ЛВП - a-глобулинов:
Определение липопротеинового спектра плазмы крови применяется в медицине для диагностики атеросклероза. Все эти липопротеины отличаются по своей функции. 1. Хиломикроны (ХМ) - образуются в клетках кишечника, их функция: перенос экзогенного жира из кишечника в ткани (в основном - в жировую ткань), а также - транспорт экзогенного холестерина из кишечника в печень. 2. Липопротеины Очень Низкой Плотности (ЛОНП) - образуются в печени, их роль: транспорт эндогенного жира, синтезированного в печени из углеводов, в жировую ткань. 3. Липопротеины Низкой Плотности (ЛНП) - образуются в кровеносном русле из ЛОНП через стадию образования Липопротеинов Промежуточной Плотности (ЛПП). Их роль: транспорт эндогенного холестерина в ткани. 4. Липопротеины Высокой Плотности (ЛВП) - образуются в печени, основная роль - транспорт холестерина из тканей в печень, то есть удаление холестерина из тканей, а дальше холестерин выводится с желчью. При определении содержания в крови липопротеинов различной плотности их обычно разделяют методом электрофореза. При этом ХМ остаются на старте, ЛОНП оказываются во фракции преb-глобулинов, ЛНП и ЛПП находят во фракции b-глобулинов, а ЛВП - a2-глобулинов. Если в крови повышено содержание b-глобулинов (ЛНП) - это означает, что холестерин откладывается в тканях (развивается атеросклероз). РОЛЬ АПОБЕЛКОВ В СОСТАВЕ ЛИПОПРОТЕИНОВ Белковые компоненты липопротеинов особенно важны. Они называются " аполипопротеинами" и обозначаются латинскими буквами (апоА, апоВ, апоС и так до апоG). ФУНКЦИИ АПОБЕЛКОВ. 1. Апобелки выполняют функцию эмульгаторов, потому что являются амфифильными веществами. 2. Некоторые из аполипопротеинов являются регуляторами активности ферментов липидного обмена. 3. Могут обладать собственной ферментативной активностью. 4. Могут выступать в качестве лигандов клеточных рецепторов для липопротеинов. 5. Многие апобелки осуществляют транспорт липидов из одного липопротеина в другой. ОБМЕН ЖИРА (ТРИАЦИЛГЛИЦЕРИНА) Химическое название жиров - ацилглицерины, то есть жиры. Это сложные эфиры глицерина и высших жирных кислот. " Ацил-" - это означает " остаток жирных кислот" (не путать с " ацетил-" - остатком уксусной кислоты). В зависимости от количества ацильных радикалов жиры разделяются на моно-, ди- и триглицериды. Если в составе молекулы 2 радикала жирных кислот, то жир называется ДИАЦИЛГЛИЦЕРИНОМ. Если в составе молекулы 1 радикал жирных кислот, то жир называется МОНОАЦИЛГЛИЦЕРИНОМ. В организме человека и животных преобладают ТРИАЦИЛГЛИЦЕРИНЫ (содержат три радикала жирных кислот).
Свойства жира определяются составом жирных кислот. Функции жира: 1. ЭНЕРГЕТИЧЕСКАЯ. В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород – группы “–CHOH-“; у жира имеются длинные углеводородные радикалы, в которых преобладают группы “-CH2-“ - в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ. Калорийность углеводов и белков: ~ 4 ккал/грамм. Калорийность жира: ~ 9 ккал/грамм. Преимуществом жира как энергетического резерва, в отличие от углеводов, является гидрофобность – он не связан с водой. Это обеспечивает компактность жировых запасов - они хранятся в безводной форме, занимая малый объем. В среднем, у человека запас чистых триацилглицеринов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме – примерно 400 гр.; при голодании этого количества не хватает даже на одни сутки. 2.ЗАЩИТНАЯ. Жировая ткань: а) защищает органы от механических повреждений. б) участвует в терморегуляции. Образование запасов жира в организме человека и некоторых животных рассматривается как приспособление к нерегулярному питанию и к обитанию в холодной среде. Особенно большой запас жира у животных, впадающих в длительную спячку (медведи, сурки) и приспособленных к обитанию в условиях холода (моржи, тюлени). У плода жир практически отсутствует, и появляется только перед рождением. ИСТОЧНИКИ ЖИРА В ОРГАНИЗМЕ: 1. Пищевой жир ( экзо генный) 2. Эндо генный жир, синтезируется в печени из углеводов. ПИЩЕВОЙ ЖИР. Переваривание экзогенного жира обязательно требует предварительного эмульгирования. Некоторые пищевые жиры поступают в организм уже в эмульгированной форме, например молочный жир. Для остальных необходимо эмульгирование с помощью специальных веществ - эмульгаторов (детергентов). Эмульгаторы – вещества амфифильной природы. Они снижают поверхностное натяжение и стабилизируют эмульсию Общее в строении эмульгаторов: наличие гидрофильных и гидрофобных участков. Гидрофильным участком молекула эмульгатора растворяется в воде, гидрофобным - в жире. Благодаря этому создается большая площадь контакта жира с водной фазой, в которой находится фермент. Белки могут выступать в роли эмульгаторов. Грудным детям эмульгаторы не нужны: они получают уже эмульгированный жир молока. В организме человека эмульгаторами являются ЖЕЛЧНЫЕ КИСЛОТЫ. Это вещества стероидной природы. Синтезируются в печени из холестерина путем окисления по монооксигеназному типу в две первичные желчные кислоты: ХОЛЕВУЮ и ХЕНОДЕЗОКСИХОЛЕВУЮ, которые затем связываются с аминокислотными остатками глицина и таурина. Так образуются конъюгированные желчные кислоты - ГЛИКОХОЛЕВАЯ (в которой гидрофильный участок представлен остатком глицина) и ТАУРОХОЛЕВАЯ (в ней гидрофильный участок представлен таурином). Гидрофобным компонентом всех желчных кислот является производное холестерина. Образуются и другие желчные кислоты – их разнообразие достаточно велико. В составе желчи желчные кислоты поступают в 12-перстную кишку и аллостерически активируют панкреатическую липазу. Собственно переваривание жиров – это гидролиз сложноэфирных связей. Существует три фермента: 1. Язычная липаза. Вырабатывается клетками слизистой оболочки задней части языка. Действие этого фермента проявляется только в желудке (раньше считали, что это – желудочная липаза). Язычная липаза может переваривать уже эмульгированный жир. Ее pH-оптимум – 4-5. Поэтому в желудке взрослого человека язычная липаза неактивна. Реально жиры перевариваются язычной липазой только у младенцев. У взрослых людей переваривание жира идет только в кишечнике по схеме: «выделение желчи Þ эмульгирование жира Þ действие панкреатической липазы». 2. Панкреатическая липаза. Сам по себе этот фермент обладает очень низкой активностью. Но в поджелудочной железе вырабатывается белок, который, попадая в кишечник, способен активировать панкреатическую липазу. Название этого белка – «колипаза». Колипаза вырабатывается в виде неактивного предшественника – проколипазы, который активируется трипсином в кишечнике. Колипаза не является классическим активатором, она лишь связывает субстрат и приближает его к активному центру липазы.
Образовавшиеся жирные кислоты и моноацилглицерины могут всасываться в кишечную стенку. 3. Эстераза липидов. Под действием этого фермента ча 4. сть моноацилглицеринов может подвергаться гидролизу с образованием глицерина и жирных кислот. Таким образом, продуктами переваривания жира являются глицерин, жирные кислоты и моноацилглицерины. Всасываются продукты переваривания путем предварительного образования смешанных МИЦЕЛЛ с желчными кислотами. Итак, желчные кислоты выполняют 2 функции: эмульгирование жира и всасывание жирных кислот. Мицеллы попадают в энтероциты. Там из компонентов мицелл снова образуются триацилглицерины, а желчные кислоты по системе воротной вены возвращаются в печень, и могут снова поступать в желчь. Этот процесс называется рециркуляцией желчных кислот. Процесс синтеза жира в энтероцитах из компонентов мицелл называется РЕСИНТЕЗОМ жира. В процессе ресинтеза происходит образование жиров, близких по составу к жирам организма. Затем из ресинтезированного жира, других липидов и апобелков формируются липопротеиновые частицы: ХИЛОМИКРОНЫ. Хиломикрон построен так же, как и остальные липопротеины (смотрите стр.2). Это небольшая жировая капля: в центре ее находятся триацилглицерины, являющиеся преобладающим компонентом частицы и составляет 80% массы хиломикрона. По периферии располагаются слои фосфолипидов (8% массы) и слои апобелков (2% массы), два из которых – А и В48 синтезируются на рибосомах энтероцита, которые чередуются. Остальные 10% массы приходятся на холестерин и его эфиры. Поверхность хиломикрона гидрофильна: гидрофильные части белков и фосфолипидов находятся на поверхности частицы. Размеры хиломикрона настолько велики, что он не может пройти через поры, имеющиеся в стенках кровеносных капилляров, путем экзоцитоза. Поэтому путем экзоцитоза хиломикроны поступают в лимфу. Через нее они попадают в большой круг кровообращения, минуя печень. После употребления в пищу жира в крови наблюдается повышенное содержание хиломикронов. В кровеносном русле происходит перенос на хиломикроны ещё двух апобелков: " С" и " Е". Стенки капилляров жировой, мышечной и других клеток, а также мембраны таких клеток содержат фермент – липопротеинлипазу. Он гидролизует триацилглицерины хиломикрона. АпоС является мощным активатором липопротеинлипазы.
Поэтому этого взаимодействия количество триацилглицеринов в хиломикроне снижается, и он теряет апобелок " С", а апоЕ при этом становится хорошим лигандом для рецепторов печени. Масса хиломикрона уменьшается. Это приводит к изменению его конформации, он превращается в «остаточный хиломикрон». Остаточный хиломикрон взаимодействует с рецепторами печени и поглощается гепатоцитами путем эндоцитоза. Печень в составе остаточного хиломикрона получает пищевой (экзогенный) холестерин. Следовательно, функциями хиломикронов являются: 1) Доставка пищевого (экзогенного) жира из кишечника в другие ткани (главным образом в жировую ткань). 2) Транспорт экзогенного холестерина из кишечника в печень. Поэтому хиломикроны - это транспортная форма экзогенного жира и экзогенного холестерина. В жировой ткани из продуктов гидролиза триацилглицеринов снова происходит ресинтез жира (второй), и он депонируется там, пока не будет востребован. ЛИПОГЕНЕЗ. АТФ необходим для синтеза активных форм субстратов, используемых в процессе синтеза жира. Для синтеза нейтрального жира необходим глицерин в активной форме - глицерол-3-фосфат (фосфоглицерин). Глицерол-3-фосфат может быть получен двумя способами: 1. Путем активации глицерина с помощью глицеринкиназы.
2. Путем восстановления фосфодиоксиацетона, полученного при распаде глюкозы.
Кроме глицерина, для синтеза нейтрального жира необходимы жирные кислоты в активной форме. Активная форма любой жирной кислоты – Ацил-КоА. Образуется при участии фермента ацил-КоА-синтазы.
Здесь наблюдается глубокий распад АТФ до АМФ. АМФ не может вступить в окислительное фосфорилирование. Поэтому существует реакция: АТФ + АМФ Þ 2АДФ. Поэтому затраты на активацию молекулы жирной кислоты эквивалентны затрате двух АТФ. Следующим этапом на пути синтеза жира является реакция образования фосфатидной кислоты:
Реакция катализируется ключевым ферментом липогенеза – глицерол-3-фосфатацилтрансферазой. Для этого фермента нет аллостерических эффекторов, но обнаружен адипсин (ацилстимулирующий белок), который облегчает взаимодействие Ацил-КоА с ферментом. Адипсин является продуктом протеолиза одного из компонентов системы комплемента. Относится к гормонам местного действия, так как вырабатывается в жировой ткани и действует там же. Две последующие реакции являются завершающими в синтезе триацилглицерина.
Реакции синтеза не зависят от того, каково происхождение веществ – участников реакций. Жир может синтезироваться как из продуктов распада жира, так и из углеводов. Синтез эндогенного жира из углеводов протекает в печени и в жировой ткани. Ацил-КоА синтезируется из Ацетил-КоА. ГБФ-путь распада углеводов обеспечивает синтез энергией. Образование Ацетил-КоА происходит в матриксе митохондрий. Синтез жирных кислот идет в цитоплазме на мембранах эндоплазматического ретикулума путем постепенного удлинения ацетил-КоА на 2 углеродных атома за каждый цикл. Удлинение высших жирных кислот, содержащих более 16 углеродных атомов, идёт путём реакций, обратных b-окислению (О b-окислении смотрите ниже, раздел «Катаболизм жирных кислот»). Однако реакции синтеза жирных кислот до 16 углеродных атомов принципиально отличаются от реакций, обратных b-окислению. Они протекают обходным обратным путём. Отличия реакций синтеза высших жирных кислот от обратных бета-окислению: 1. b-окисление протекает в митохондриях, а синтез жирных кислот протекает в цитоплазме на мембранах эндоплазматического ретикулума. Но образуется Ацетил-КоА в митохондриях, а через мембраны сам проходить не может. Поэтому существуют механизмы транспорта Ацетил-КоА из митохондрий в цитоплазму.
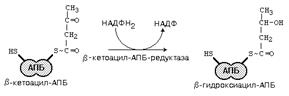
Ключевыми ферментами ЦТК являются цитратсинтаза и изоцитратдегидрогеназа. Основные аллостерические регуляторы этих ферментов - это АТФ и АДФ. Если в клетке много АТФ, то АТФ выступает как ингибитор этих ключевых ферментов. Однако изоцитратдегидрогеназа угнетается АТФ сильнее, чем цитратсинтетаза. Это приводит к накоплению цитрата и изоцитрата в матриксе митохондрии. При накоплении цитрат выходит из митохондрии в цитоплазму. В цитоплазме есть фермент цитратлиаза. Этот фермент расщепляет цитрат на ЩУК и Ацетил-КоА. Условием для выхода Ацетил-КоА из митохондрии в цитоплазму является хорошее обеспечение клетки АТФ. Если АТФ в клетке мало, то Ацетил-КоА расщепляется до СО2 и Н2О. 2. В ходе b-окисления промежуточные продукты связаны с HS-КоА, а при синтезе жирных кислот промежуточные продукты связаны с особым ацил-переносящим белком (АПБ). Это сложный белок. Его небелковая часть похожа по строению на КоА. Небелковая часть АПБ состоит из тиоэтиламина, пантотеновой кислоты (витамин В3) и фосфата. 3. При b-окислении в качестве окислителя используются НАД и ФАД. При синтезе ЖК нужен восстановитель - используется НАДФ*Н2. В клетке существует 2 основных источника НАДФ*Н2 для синтеза жирных кислот: а) ГМФ-путь распада углеводов б) В реакции окисления малата (смотрите рисунок).
Эта реакция протекает в цитоплазме и катализируется ферментом МАЛАТДЕГИДРОГЕНАЗОЙ ДЕКАРБОКСИЛИРУЮЩЕЙ, тривиальное название которого – ЯБЛОЧНЫЙ ФЕРМЕНТ. 4. В ходе синтеза жирных кислот в каждом цикле удлинения используется не сам Ацетил-КоА, а его производное - малонил-КоА (при b-окислении каждый цикл укорочения приводит к образованию Ацетил-КоА).
Эту реакцию катализирует фермент АЦЕТИЛ-КоА-КАРБОКСИЛАЗА. Это ключевой фермент в мультиферментной системе синтеза ЖК. Этот фермент регулируется по типу отрицательной обратной связи. Ингибитором является продукт синтеза: ацил-КоА с длинной цепью (n=16) - пальмитоил-КоА. Активатором является цитрат. В состав небелковой части этого фермента входит витамин H (биотин). Далее происходит поэтапное удлинение молекулы Ац-КоА на 2 углеродных атома за каждый этап за счет малонил-КоА. В процессе удлинения малонил-КоА теряет СО2. После образования малонил-КоА основные реакции синтеза жирных кислот катализируются одним ферментом - синтетазой жирных кислот (фиксирован на мембранах эндоплазматического ретикулума). Синтетаза жирных кислот содержит 7 активных центров. Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту).
РЕАКЦИИ СИНТЕЗА ЖИРНЫХ КИСЛОТ.
После этого ацил-АПБ вступает в новый цикл синтеза. К свободной SH-группе ацилпереносящего белка присоединяется новая молекула малонил-КоА. Затем происходит отщепление ацильного остатка, и он переносится на малонильный остаток с одновременным декарбоксилированием, и цикл реакций повторяется. Таким образом, углеводородная цепочка будущей жирной кислоты постепенно растет (за каждый цикл – на два углеродных атома). Это происходит до момента, пока она не удлинится до 16 углеродных атомов (в случае синтеза пальмитиновой кислоты) или более (синтез других жирных кислот). Вслед за этим происходит тиолиз, и образуется в готовом виде активная форма жирной кислоты – ацил-КоА. УСЛОВИЯ ДЛЯ ПРОТЕКАНИЯ СИНТЕЗА ВЫСШИХ ЖИРНЫХ КИСЛОТ: 1) Поступление углеводов, при окислении которых образуются необходимые субстраты и НАДФН2. 2) Высокий энергетический заряд клетки – высокое содержание АТФ, которое обеспечивает выход цитрата из митохондрий в цитоплазму. Обмен углеводов и обмен жиров очень тесно связаны. Углеводы легко могут превращаться в жиры, а вот превращение жиров в углеводы невозможно. Жиры не могут превращаться в углеводы, так как Ацетил-КоА не может превращаться в пируват. Обмен жиров и углеводов объединяется как энергетический обмен, который находится под контролем гормонов. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 502; Нарушение авторского права страницы