|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Билет 13. Множественный аллелизм. Школа Серебровского по ступенчатому аллелизму. Функциональный тест.
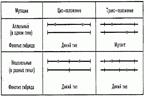
Одним из первых доказательств сложности гена явилось обнаружение явления множественного аллелизма, свидетельствующего о большей функциональной лабильности гена, чем это думали раньше. В 1929 - 1930 гг. в нашей стране в работах А. С. Серебровского и его молодых сотрудников - Н. П. Дубинина, Б. Н. Сидорова и других - была впервые экспериментально показана функциональная сложность гена. Авторы исследовали у дрозофилы серию множественных аллелей фокуса scute, локализованного в нулевой точке половой хромосомы. Мутации этого локуса st, scs и другие обусловливают редукцию разных щетинок на теле мухи. При скрещивании особей, как правило, отсутствовали лишь те щетинки, которые были редуцированы у обеих гомозигот. Так, например, если одна мутация sc1 вызывала редукцию щетинок АВС, а другая — редукцию щетинок ВСР, то у гетерозиготы, отсутствовали щетинки В и С, а щетинки А и D имелись. Создавалось впечатление, что в данном случае речь идет о частичной гетерозиготности, когда части мутантных аллелей, которые обусловливают одинаковый фенотипический эффект, оказываются в гомозиготном состоянии. Поэтому описанное явление получило название ступенчатого аллелизма. Псевдоаллелизм. Представление о гене как единице, далее не делимой кроссинговером, подразумевало, что при гаметогенезе у компаундов, т.е. зигот, несущих две аллели одной серии (а1│ │ а2) могут образовываться гаметы только двух типов – а1 и а2. При возвратном скрещивании таких особей с любой из родительских форм возможно появление только мутантных форм. Однако если выборку увеличить, например, до l00 тыс. и более особей, то в ней окажутся и потомки дикого типа. Такие особи могли появиться только при двух условиях: мутация затрагивает часть гена дикого типа и между частями гена может происходить кроссинговер. Явление это было открыто Е. Льюисом и другими при изучении ряда генов у дрозофилы. Существование такого явления противоречило представлению о гене как единице, далее неделимой при кроссинговере. Однако трудно было сразу отказаться от традиционных представлений, и об аллелях, делимых при кроссинговере, стали говорить как о псевдоаллеллх. Псевдоаллелизм распространен весьма широко. Он был продемонстрирован на разнообразных организмах: аспергилле, нейроспоре, дрожжах, хлопчатнике, кукурузе, шелкопряде, дрозофиле, голубях, мышах, норках и многих других объектах. Исходя из того, что ген, согласно современным данным, представляет собой сложную линейную структуру, а мутации могут затрагивать различные его участки, были сделаны попытки модернизировать моргановский функциональный критерий аллелизма. Цис-транс-тест на аллелизм. Льюис предложил цис-транс-тест на аллелизм. Смысл этого теста сводится к тому, что при скрещивании двух мутантных особей возникает зигота с транс-конфигурацисй этих мутаций. Для проведения Ц.-т.-т. исследуемые мутации сочетают в транс- и цис-положениях: в первом случае скрещивают особи, несущие по одной анализируемой мутации, во втором — скрещивают особь, несущую обе мутации, с особью дикого (нормального) типа. Если мутации, сочетаемые в транс-положении (транс-тест, или функциональный тест на аллелизм), принадлежат разным генам, то гибридный организм автоматически получает и по неповрежденной копии каждого гена. В этом случае рецессивные мутации не проявляются и гибрид имеет нормальный фенотип (мутации комплементарны; см. Комплементация). Если сочетаемые мутации принадлежат одному гену, то в гибриде обе копии данного гена повреждены и обнаруживается мутантный фенотип (мутации некомплементарны). Усовершенствование транс-теста, предложенное Э. Льюисом, состоит в исследовании сочетаемых мутаций также в цис-положении для исключения артефактов за счёт взаимодействия генов на уровне генных продуктов. Если фенотип гибрида при Ц.-т.-т. одинаков в цис- и транс-положениях (нет цис-транс-эффекта), то исследуемые мутации принадлежат разным генам. Если же фенотип гибрида в цис- и транс-положениях различен (есть цис-транс-эффект), то мутации принадлежат одному гену. В практике генетического анализа по-прежнему широко применяется транстест для выявления принадлежности мутаций одному гену (цистрону).
Билет 14. Исследование гена на примере фага Т-4. Физ и хим размеры карты. Ген как ед. ф-ии(цистрон) Классической работой, показавшей пределы мутационной и рекомбинационной делимости гена, стало исследование фага Т4, инфицирующего бактерию E. coli, проведенное в конце 50-х годов С. Бензером. Он изучал мутантные формы r II. Взаимоотношения мутантов r II c различными штаммами E. сoli создают возможности для: 1. выращивания только ревентантов и рекомбинантов r+ (на штамме E. coli К 12(λ ) ), 2. одновременного исследования мутантов r II и фагов дикого типа r+ (на штамме E. coli В), 3. размножения r II и фагов дикого типа r+ (на штамме E. coli S). при заражении бактериальной клетки двумя фаговыми частицами между их геномами возникают связи, как между гомологичными хромосомами диплоида. При этом признаки фага дикого типа доминируют – образуются стерильные пятна стандартного типа на штаммах В и К 12 (λ ). Т.о., возможно проведение функционального теста на аллелизм. Локус r II был разбит на два функционально самостоятельных участка гена, или как их назвал Бензер, цистрона А и В. Функционально самостоятельные участки гена были названы цистронами. Цистрон представляет собой участок хромосомы, мутация в пределах которого обнаруживается в транс-положении. Понятие «ген» в функциональном отношении шире понятия «цистрон». Цистрон — участок ДНК, кодирующий первичную структуру одной полипептидной цепи или одной молекулы РНК (р-РНК или т-РНК). Ген же — участок ДНК, кодирующий первичную структуру одной молекулы белка, независимо от того, состоит он из одной субъединицы (в этом смысле понятия «ген» и «цистрон» совпадают) или из нескольких (И. П. Ашмарин, 1977)'. Бензер попытался дать конкретное выражение размеров цистрона. Исходя из того, что вся ДНК фага содержит 4.10~ нуклеотидов и 40 Щ из них составляют генетический материал, он установил, что каждая функциональная единица включает несколько сотен пар нуклеотидов: ген гПЛ — 800, rllB — 1500. Опыты Бензера позволили установить также наименьшую единицу рекомбинации. Оказалось, что даже соседние основания в молекуле ДНК могут отделяться друг от друга путем кроссинговера. Таким образом, ген представляет собой весьма сложную структуру. По словам Дж. Уотсона, «ген — это дискретная область хромосомы, ответственная за образование определенного клеточного продукта; он состоит из ряда линейно расположенных единиц, потенциально способных к изменению (мутирующие участки); каждый такой участок может существовать в нескольких альтернативных формах и между разными участками может происходить кроссинговер». В ходе редупликации фаговых геномов после инфекции бактериального штамма двумя мутантами rll могут образовываться рекомбинанты дикого типа. Частота появления рекомбинантов в таких скрещиваниях характеризует расстояние между мутантными точками в геноме бактериофага. Используя селективные возможности перечисленных штаммов Е. coli, С. Бензер инфицировал клетки Е. соli S парой мутантов, высевал фаголизат на штаммы К и S, а затем сравнивал появление негативных колоний на этих штаммах. При попарном исследовании способности к рекомбинации трех различных мутантов rII, получаемые значения частот рекомбинации могут быть использованы для линейного расположения мутаций. К 1962 г. в пределах участков А и В локуса rll было картировано более 2000 мутаций.
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 127; Нарушение авторского права страницы