|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Азнообразие свойств нервных центров. ⇐ ПредыдущаяСтр 4 из 4
Свойства эти следующие: 1. Одностороннее проведение в нервных центрах можно доказать при раздражении передних корешков и отведении потенциалов от задних. В этом случае осциллограф не зарегистрирует импульсов. Если поменять электроды - импульсы будут поступать нормально. 2. Задержка проведения в синапсах. По рефлекторной дуге проведение возбуждения происходит медленнее, чем по нервному волокну. Это определяется тем, что в одном синапсе переход медиатора к постсинаптической мембране происходит за 0, 3-0, 5 мсек. (т.н. синаптическая задержка). Чем больше синапсов в рефлекторной дуге, тем больше время рефлекса, т.е. интервал от начала раздражения до начала деятельности. С учетом синаптической задержки проведение раздражения через один синапс требует около 1, 5-2 мсек. У человека наименьшую продолжительность имеет время сухожильных рефлексов (оно равно 20-24 мсек). У мигательного рефлекса оно больше - 50-200 мсек. Время рефлекса складывается из: а) времени возбуждения рецепторов; б) времени проведения возбуждения по центростремительным нервам; в) времени передачи возбуждения в центре через синапсы; г) времени проведения возбуждения по центробежным нервам; д) времени передачи возбуждения на рабочий орган и латентного периода его деятельности. Время " в" носит название центрального времени рефлекса.
3. Суммация возбуждений. Впервые Сеченов показал, что в целостном организме рефлекторный акт может осуществляться при действии подпороговых стимулов, если они действуют на рецепторное поле достаточно часто. Такое явление получило название временной (последовательной) суммацией. Пример - рефлекс чесания у собаки можно вызвать, если подать в одну точку подпороговые стимулы с частотой 18 гц. Суммация подпороговых стимулов можно получить и тогда, когда они прикладываются на разные точки кожи, но одновременно - это пространственная суммация. В основе этих явлений лежит процесс суммации возбуждающих постсинаптических потенциалов на теле и дендритах нейронов. При этом происходит накопление медиатора в синаптической щели. В естественных условиях оба вида суммации сосуществуют.
4. Центральное облегчение. Возникновение временной и особенно пространственной суммации способствуют и особенности организации синаптического аппарата в нервных центрах. Каждый аксон, поступая в ЦНС, ветвится и образует синапсы на большой группе нейронов (нейронный пул, или нейронная популяция). В такой группе принято условно различать центральную (пороговую) зону, и периферическую (подпороговую) кайму. Нейроны, находящиеся в центральной зоне, получают от каждого рецепторного нейрона достаточное количество синаптических окончаний для того, чтобы ответить разрядом ПД на приходящие импульсы. На нейронах же подпороговой каймы каждый аксон образует лишь небольшое число синапсов, возбуждение которых не способно возбудить нейрон. Нервные центры состоят из большого числа нейронных групп, причем отдельные нейроны могут входить в разные нейронные пулы. Это объясняется тем, что на одних и тех же нейронах оканчиваются разные афферентные волокна. При совместном раздражении этих афферентных волокон возбуждающие постсинаптические потенциалы в нейронах подпороговой каймы суммируются друг с другом и достигают критической величины. В результате в процесс возбуждения оказываются вовлеченными и клетки периферической каймы. При этом сила рефлекторной реакции суммарного раздражения нескольких " входов" в центр оказывается больше арифметической суммы раздельных раздражений. Этот эффект и носит название центрального облегчения.
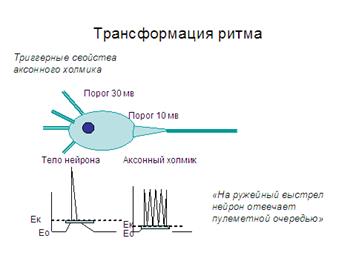
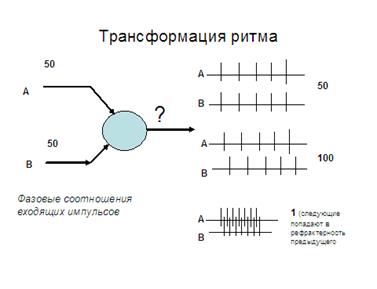
6. Трансформация ритма возбуждений. Частота и ритм импульсов, поступающих к нервным центрам, и посылаемых ими на периферию, могут не совпадать. Это явление носит название трансформации. В ряде случаев на одиночный импульс, приложенный к афферентному волокну, мотонейрон отвечает серией импульсов. Образно говоря, в ответ на одиночный выстрел нервная клетка отвечает очередью. Чаще это бывает при длительном постсинаптическом потенциале и зависит от триггерных свойств аксонного холмика. Другой механизм трансформации связан с эффектами сложения фаз двух или более волн возбуждения на нейроне - тут возможны эффекты как увеличения, так и снижения частоты выходящих из центра стимулов.
7. Последействие. Рефлекторные акты, в отличие от потенциалов действия, заканчиваются не одновременно с прекращением вызвавшего их раздражения, а через некоторый, иногда сравнительно длинный период времени. Продолжительность последействия может во много раз превышать продолжительность раздражения. Последействие обычно бывает больше при сильном и длительном раздражении. Имеются два основных механизма, обусловливающих эффект последействия. Первый связан с суммацией следовой деполяризации мембраны при частых раздражениях (посттетаническая потенциация), когда нервная клетка продолжает давать разряды импульсов, несмотря на то, что кончилась серия раздражений. Второй механизм связывает последействие с циркуляцией нервных импульсов по замкнутым нейронным сетям рефлекторного центра. 8. Утомление нервных центров. В отличие от нервных волокон нервные центры легко утомляемы. Утомление нервного центра проявляется в постепенном снижении и в конечном итоге полном прекращении рефлекторного ответа при продолжительном раздражении афферентных нервных волокон. Если после этого приложить раздражение в эфферентному волокну - эффект возникает вновь. Утомление в нервных центрах связано прежде всего с нарушением передачи возбуждения в межнейронных синапсах. Такое нарушение зависит от уменьшения запасов синтезированного медиатора, уменьшением чувствительности к медиатору постсинаптической мембраны, уменьшением энергетических ресурсов нервной клетки. Не все рефлекторные акты утомляются быстро (например, мало утомляемы проприоцептивные тонические рефлексы). 9. Рефлекторный тонус нервных центров. В его поддержании участвуют как афферентные импульсы, поступающие непрерывно от периферических рецепторов в ЦНС, так и различные гуморальные раздражители (гормоны, углекислота, и др.) 10. Высокая чувствительность к гипоксии. Показано, что 100 г. нервной ткани в единицу времени потребляет кислорода в 22 раза больше, чем 100 г. мышечной ткани. Поэтому нервные центры очень чувствительны к его недостатку. При этом чем выше центр, тем более страдает он от гипоксии. Для коры мозга 5-6 минут достаточно, чтобы без кислорода произошли необратимые изменения, клетки ствола мозга выдерживают 15-20 минут полного прекращения кровообращения, а клетки спинного мозга - 20-30 минут. При гипотермии, когда снижается обмен веществ, ЦНС дольше переносит гипоксию. 11. Избирательная чувствительность к химическим веществам. Объясняется особенностями обменных процессов и позволяет находить фармпрепараты направленного действия. Такие свойства как однонаправленность проведения, замедление проведения нервных импульсов, и суммация непосредственно вытекают из свойств отдельных нейронов. Усвоение и трансформация ритма приходящих в центр импульсов проявляются в способности центра возбуждаться или тормозиться в соответствии с ритмом поступающих сигналов. Это свойство имеет значение для взаимодействия нервных центров при организации поведенческих актов. Кроме того, нервный центр способен превращать ритм приходящих импульсов в свой собственный. ^ 2. Понятие о рефлексе. Выдвинутая в XVIII в. идея о рефлекторном, ответном характере нервных процессов, обеспечивающих связь человека с внешним миром, приобрела интенсивное развитие в XIX – XX вв. на основе новых неопровержимых экспериментальных данных, полученных строгими научными методами. Она началось с объяснения отдельных простейших движений человека, а затем была распространена на всю высшую нервную деятельность и переформулирована в её рефлекторный принцип. «Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы», – писал И.М. Сеченов в своей работе «Рефлексы головного мозга». Рефлекс – ответ организма на внешнее воздействие. Это одна из функциональных единиц высшей нервной деятельности. Её действие обеспечивается возникновением и распространением возбуждения от рецепторов по центростремительному – афферентному (сенсорному) – нервному пути к мозговым центрам, далее по центральному нервному пути, «переадресующему» поступившее в мозг возбуждение от вызванного им очага к области, участку мозга, представляющим в нём какой-то орган, и, наконец, от него по центробежному – эфферентному (исполнительному) – пути от них к органам, осуществляющим соответствующее ответное движение, сокращение, выделение и пр. Этот трёхзвенный нервный путь упрощённо изображается в виде «дуги рефлекса».
^ 3. Условный рефлекс и его формирование. В реальной жизнедеятельности человека значительно больше нестандартных ситуаций и факторов, чем стандартных, а поэтому безусловные рефлексы не могут обеспечивать правильное поведение во всём многообразии жизни. Зато это в возможностях условных рефлексов. Они не врожденны, и у них нет готового с рождения нервного пути, в начале которого раздражение, а в конце обязательный эффекторный ответ. Появляющийся на свет ребёнок имеет лишь набор разных нервных элементов, нужных для нестандартной реакции: афферентных, центральных, эффекторных, но не соединенных в одну цепь «дуг рефлекса». Это нейронный набор представляет собой потенциал для прижизненного соединения их в любых вариантах и образования новых рефлекторных путей у растущего и развивающегося человека в опыте его встреч с нестандартными ситуациями. Мозг способен использовать этот потенциал и формировать новые способы реагирования на нестандартные условия, т.е. условные рефлексы. Большой вклад в их исследование внёс И.П. Павлов, используя разработанную им оригинальную методику. Она использовалась при изучении слюноотделительных рефлексов у собак. Животное подвергалось операции: один из слюноотделительных протоков выводился из ротовой полости на поверхность кожи, к нему прикреплялась пробирка, в которую выделялась слюна. Это давало возможность точно определять: когда, в каком количестве, с какой интенсивностью у собаки выделяется слюна. Далее животное помещали в специальную звуконепроницаемую камеру, отдельную от экспериментатора, фиксировали ремнями. Для вызова рефлекторного слюноотделения применяли пищевой раздражитель. В камере находились приборы для вызова разнообразных раздражений: звонок, лампочки, кожные касалки и др. процедура выработки нового рефлекса сводилась к следующему:
1. 2. 3. 4. Регистрация слюноотделения производилась по шкале, на которой укреплена трубка: слюна, поступающая из резервуарчика на щеке собаки в баночку Б, вытесняла из неё воздух, который давил на жидкость в трубке А. Общий механизм образования условных рефлексов можно изобразить схематично. Вначале используется индифферентный раздражитель – безразличный индивиду, особи по прошлому опыту, а поэтому не имеющий при первом воздействии значения и не вызывающий никакой другой реакции, кроме ориентировочной. Он вызывает центростремительное возбуждение, которое 6 6 образует относительно слабый очаг возбуждения в мозговых центрах и коре больших полушарий. Вслед за этим приводится в действие имеющий значение раздражитель (у И.П. Павлова – пища), который тоже вызывает очаг возбуждения, но несравненно более сильный Классификация рефлексов
Основным механизмом деятельности ЦНС является рефлекс как ответная реакция организма на действия раздражителя, осуществляемая с участием ЦНС. В переводе с латинского языка это слово означает «отражение». Впервые данный термин был применен французским философом Р.Декартом для характеристики реакций организма в ответ на раздражение органов чувств. Он первым высказал мысль о том, что все проявления эффекторной активности организма вызываются вполне реальными физическими факторами. Иначе говоря, Декарт теоретически показал, что всякое действие имеет вполне реальную физическую причину. После Р.Декарта представление о рефлексе было развито чешским исследователем Й.Прохазкой, который развил учение об отражательных действиях. Важной вехой в развитии рефлекторной теории стала книга «Рефлексы головного мозга» И.М.Сеченова. Он утверждал, что все акты бессознательной и сознательной жизни по природе происхождения являются рефлексами. Эту мысль экспериментально подтвердил И.П.Павлов, разработав метод условных рефлексов. Он доказал, что высшая нервная деятельность, так же как и низшая, является рефлекторной. Морфологическим субстратом рефлекса является рефлекторная дуга – совокупность морфологических структур, которые обеспечивают осуществление рефлекса. Иными словами, рефлекторная дуга является путем, по которому проходит возбуждение при осуществлении рефлекса. Согласно рефлекторной теории И.П.Павлова, в рефлекторной дуге следует различать пять структурных элементов: рецептор, афферентный путь, центральное звено, эфферентный путь и эффектор. Рецептор — это чувствительное нервное окончание, воспринимающее раздражение. В рецепторах энергия раздражителя превращается в энергию нервного импульса. Различают: 1) экстерорецепторы — возбуждаются под влиянием раздражений из окружающей среды (рецепторы, кожи, глаза, внутреннего уха, слизистой оболочки носа и ротовой полости); 2) интерорецепторы — воспринимают раздражения из внутренней среды организма (рецепторы внутренних органов, сосудов); 3) проприорецепторы — реагируют на изменение положения отдельных частей тела в пространстве (рецепторы мышц, сухожилий, связок, суставных сумок). Афферентный нервный путь представлен отростками рецепторных нейронов, несущих возбуждения в центральную нервную систему. Рефлекторный центр состоит из группы нейронов, расположенных на различных уровнях центральной нервной системы и передающих нервные импульсы с афферентного на эфферентный нервный путь. Эфферентный нервный путь проводит нервные импульсы от ЦНС к эффектору. Афферентный (чувствительный) путь – это система нервных волокон, которая обеспечивает проведение электрического импульса от рецептора к центру рефлекса. В центральном звене рефлекса происходит обработка полученной информации и формируется программа ответной реакции. Эфферентный (двигательный) путь - это система нервных волокон, которая передает импульсы от коры или ниже лежащих ядер головного мозга через спинной мозг к рабочему органу. Эффектор — исполнительный орган деятельность которого изменяется, под влиянием нервных импульсов, поступающих к нему по образованиям рефлекторной дуги. Эффекторами могут быть мышцы или железы. По современным представлениям структурной основой рефлекса является рефлекторное кольцо, которое состоит из следующих звеньев: рецептор, афферентный путь, нервный центр, эфферентный путь, эффектор, обратная афферентация. Морфологическим субстратом обратной афферентации являются рецептор и афферентное звено рефлекса. Принцип обратной афферентации введен в рефлекторную теорию П.К. Анохиным, который высказал мысль о том, что для рефлекторной деятельности необходимо наличие полезного приспособительного результата, ради которого совершается любой рефлекс. Информация о достижении или не достижении полезного приспособительного результата поступает в центр рефлекса по звену обратной связи в виде обратной афферентации, которая является обязательным компонентом рефлекторной деятельности. Существует несколько классификаций рефлексов. Так, И.М.Сеченов выделил следующие виды рефлексов: 1. невольные движения, которые заключают в себе чистые рефлексы и рефлексы с психическим элементом; 2. произвольные движения, заключающие в себе рефлексы с психическими элементами. Структуру чистого рефлекса И.М.Сеченов представил следующим образом: возбуждение чувствующего нерва → возбуждение спинномозгового центра, связывающего чувствующий нерв с двигательным нервом → возбуждение двигательного нерва → движение Чистые рефлексы по Сеченову возможны при определенных условиях: а) если спинной мозг изолирован от головного; б) если чувствительный нерв раздражается неожиданно; в) если сила раздражения значительно превосходит среднюю величину; г) во время сна; д) если внимание отвлечено от данного раздражения. В настоящее время основной глобальной классификацией является разделение всех рефлексов на безусловные и условные (по Павлову). Далее идет разделение в зависимости от существующих характеристик рефлексов. 1. Классификация по рецепторному звену. · Интероцептивные: информация, возбуждающая рецептор и, запускающая рефлекс, получена с рецепторов внутренних органов; · Экстероцептивные: информация, возбуждающая рецептор и, запускающая рефлекс, получена из внешней среды с помощью сенсорных систем; · Проприоцептивные: рефлексы, запускаемые с рецепторов мышц, сухожилий и суставов. · по центральному звену выделяют: центральные (истинные) – главное звено находится в ЦНС и периферические – центральное звено находится за пределами ЦНС. Центральные, в свою очередь делятся на спинальные и церебральные. Спинальные рефлексы делят на цервикальные, торакальные, люмбальные и сакральные. Церебральные рефлексы делят на мозжечковые, рефлексы больших полушарий и рефлексы ствола мозга. Рефлексы ствола мозга делят на бульбарные, диэнцефальные и мезенцефальные. 2. Классификация по эффекторам. · Двигательные; · Сердечные; · Сосудистые; · Секреторные. 3. Классификация, основанная одновременно на рецепторном и эффекторном звене: висцеро-висцеральные, сенсоро-висцеральные, висцеро-соматические, сомато-висцеральные. 4. Классификация рефлексов по их биологической значимости. Здесь выделяют оборонительные и защитные, половые, пищевые и питьевые, ориентировочно-исследовательские, родительские, гомеостатические. 5. Классификация рефлексов по количеству синаптических контактов или нейронов в рефлекторной дуге. · Двухнейронные, или моносинаптические, рефлекторные дуги характеризуются наличием в своем составе только двух нейронов (чувствительного и двигательного). В такой дуге участвует один синапс, который находится в ЦНС. Эти рефлексы участвуют в регуляции мышечного тонуса и позы и связаны, в первую очередь, с натяжением сухожилий. В случае двухнейронной организации дуги рефлекторные акты осуществляются без участия головного мозга, так как рефлексы стереотипны и не требуют обдумывания или сознательного решения. · Полинейронные, или полисинаптические, рефлекторные дуги обеспечивают подавляющее большинство рефлекторных актов. В них участвуют как минимум два синапса, находящиеся в ЦНС, так как в дугу включен третий нейрон – вставочный (интернейрон). Такие рефлекторные дуги позволяют организму осуществлять автоматические непроизвольные реакции, необходимые для приспособления к изменениям внешней и внутренней среды. Все рефлексы обладают определенным общим набором свойств. Проведение сигнала по рефлекторной дуге осуществляется только в одном направлении: от рецептора к центру, а от центра к эффектору. Морфо-функциональной основой этого свойства является одностороннее проведение в химических синапсах. В ответ на раздражитель реакция происходит не мгновенно, а спустя некоторое время, которое называется латентным периодом рефлекса или временем рефлекса. Латентное время рефлекса формируется как сумма: · Латентного периода возбуждения рецептора; · Времени проведения ПД по афферентному пути; · Синаптической задержки в центральном звене рефлекса; · Времени проведения ПД по эфферентному пути; · Латентного периода ответа органа-эффектора. - Чем больше синапсов в рефлекторной дуге, тем больше будет латентное время рефлекса. - Сила рефлекса растет с усилением раздражения рецепторов или с увеличением площади воздействия раздражителя. - Полисинаптическим рефлексам свойственен эффект последействия. У моносинаптических рефлексов последействия нет. Сущность эффекта заключается в продлении ответа некоторое время после того, как внешнее раздражение прекращено. В основе эффекта последействия полисинаптического рефлекса лежат следующие причины: 1. большая длительность ВПСП вставочных нейронов, которые порождают не один ПД, а серию; 2. асинхронность сигналов, достигающих мотонейронов по параллельным цепочкам нейронов разной сложности и с разными скоростями срабатывания. - Многие рефлекторные центры обладают свойством постактивационной потенциации, суть, которой заключается в усилении ответов на одиночные тестирующие сенсорные сигналы в течение некоторого времени после окончания ритмической активности. Данный эффект не зависит от количества нейронов в рефлекторном кольце и связан в первую очередь с модификацией синапса в ходе ритмического воздействия. Рефлекторная дуга.
Рефлекторная дуга или рефлекторный путь представляет собой совокупность образований, необходимых для осуществления рефлекса. В нее входит цепь соединенных посредством синапсов нейронов, которая передает нервные импульсы от возбужденных стимулом чувствительных окончаний к мышцам или секреторным железам. В рефлекторной дуге различают следующие компоненты: 1.Рецепторы-высокоспециализированные образования, способные воспринять энергию раздражителя и трансформировать ее в нервные импульсы. Все рецепторы можно подразделить на внешние или экстерорецепторы( зрительные, слуховые, вкусовые, обонятельные, осязательные) и внутренние или интерорецепторы( рецепторы внутренних органов), среди которых полезно выделить проприорецепторы, находящиеся в мышцах, сухожилиях и суставных сумках. 2.Сенсорные ( афферентные, центростремительные ) нейроны, проводящие нервные импульсы от своих дендритов в центральную нервную систему. В спинной мозг сенсорные волокна входят в составе задних корешков. 3.Интернейроны ( вставочные, контактные) находятся в центральной нервной системе, получают информацию от сенсорных нейронов, перерабатывают ее и передают эфферентным нейронам. В спинном мозгу тела вставочных нейронов находятся преимущественно в задних рогах и промежуточной области. 4.Эфферентные ( центробежные) нейроны получают информацию от интернейронов ( в исключительных случаях от сенсорных нейронов) и передают рабочим органам.Тела эфферентных нейронов расположены в центральной нервной системе, а их аксоны выходят из спинного мозга в составе передних корешков и относятся уже к периферической нервной системе: они направляются либо к мышцам, либо к внешнесекреторным железам. Управляющие скелетными мышцами двигательные нейроны спинного мозга (мотонейроны) находятся в передних рогах, а вегетативные нейроны- в боковых рогах. Для обеспкчения соматических рефлексов достаточно одного эфферентного нейрона, а для осуществления вегетативных рефлексов необходимо два: один из них располагается в центральной нервной системе, а тело другого находится в вегетативном ганглии. 5.Рабочие органы или эффекторы представляют собой либо мышцы, либо железы, поэтому рефлекторные ответы в конечном счете сводятся или к мышечным сокращениям (скелетных мышц, гладких мышц сосудов и внутренних органов, сердечной мышцы), или к выделению секретов желез (пищеварительных, потовых, бронхиальных, но не желез внутренней секреции). Благодаря химическим синапсам возбуждение по рефлекторной дуге распространяется только в одном направлении: от рецепторов к эффектору. В зависимости от количества синапсов различают полисинаптические рефлекторные дуги, в состав которых входит не менее трех нейронов (афферентный, интернейрон, эфферентный), и моносинаптические, состоящие лишь из афферентного и эфферентного нейронов. У человека моносинаптические дуги обеспечивают воспроизведение только рефлексов растяжения, регулирующих длину мышц, а все остальные рефлексы осуществляются с помощью полисинаптических рефлекторных дуг. Элементарные дуги спинальных рефлексов посредством проводящих путей взаимодействуют с высшими центрами головного мозга. Кроме того к классическим компонентам рефлекса (стимул-нервный центр-ответ) следует добавить обратную связь, т. е. механизм предоставления информации о том, удалось или нет с помощью рефлекторной реакции приспособиться к изменениям среды и насколько эффективным оказалось приспособление. |
Последнее изменение этой страницы: 2017-03-13; Просмотров: 698; Нарушение авторского права страницы
 Для упомянутых выше рефлексов оно составляет соответственно 3 мсек. и 36-180 мсек. Зная центральное время рефлекса, и учитывая, что через один синапс возбуждение проходит за 2 мсек., можно определить число синапсов в рефлекторной дуге. Например, коленный рефлекс считают моносинаптическим.
Для упомянутых выше рефлексов оно составляет соответственно 3 мсек. и 36-180 мсек. Зная центральное время рефлекса, и учитывая, что через один синапс возбуждение проходит за 2 мсек., можно определить число синапсов в рефлекторной дуге. Например, коленный рефлекс считают моносинаптическим. 
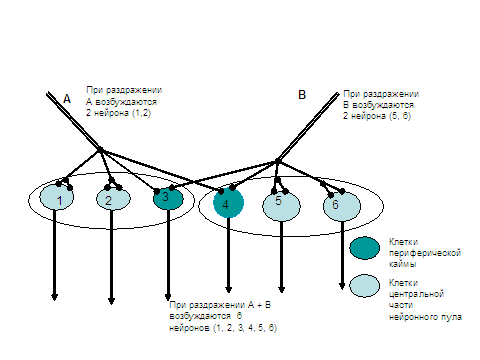
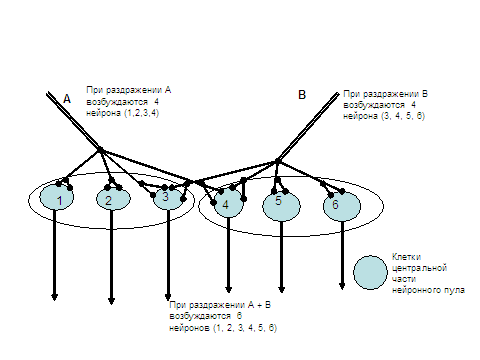 5. Центральная окклюзия (закупорка ). Может наблюдаться в деятельности нервного центра и обратный эффект, когда одновременное раздражение двух афферентных нейронов вызывает не суммацию возбуждения, а задержку, уменьшение силы раздражения. В этом случае суммарная реакция меньше арифметической суммы раздельных эффектов. Происходит это потому, что отдельные нейроны могут входить в центральные зоны разных нейронных популяций. В таком случае появление возбуждающих постсинаптических потенциалов на телах нейронов не приводят к увеличению числа возбужденных одновременно клеток. Если суммация лучше проявляется при действии слабых афферентных раздражений, то явления окклюзии хорошо выражены с случае применения сильных афферентных раздражений, каждое их которых активирует большое число нейронов. Более наглядно эти эффекты видны на схемах в таблицах.
5. Центральная окклюзия (закупорка ). Может наблюдаться в деятельности нервного центра и обратный эффект, когда одновременное раздражение двух афферентных нейронов вызывает не суммацию возбуждения, а задержку, уменьшение силы раздражения. В этом случае суммарная реакция меньше арифметической суммы раздельных эффектов. Происходит это потому, что отдельные нейроны могут входить в центральные зоны разных нейронных популяций. В таком случае появление возбуждающих постсинаптических потенциалов на телах нейронов не приводят к увеличению числа возбужденных одновременно клеток. Если суммация лучше проявляется при действии слабых афферентных раздражений, то явления окклюзии хорошо выражены с случае применения сильных афферентных раздражений, каждое их которых активирует большое число нейронов. Более наглядно эти эффекты видны на схемах в таблицах.