
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Хромосомы и их химический состав.
Митоз и его генетическая сущность. Мейоз и его генетическая сущность. Оплодотворение: случайность и избирательность при слиянии гамет. VІІ. Внеядерная наследственность. Материнская наследственность и ее причины. Наследственность, передаваемая только через мать, называется материнской. Материнская наследственность характерна лишь для тех признаков на которые влияют цитоплазма и ее органоиды. Наследование признаков через цитоплазму получило название ЦИТОПЛАЗМАТИЧЕСКОЙ, или ВНЕЯДЕРНОЙ наследственности. Причины осложняющие правильные решения роли цитоплазмы в наследственности: 1.Иногда в цитоплазме находятся инородные органический с ней не связанные включнения, чаще вирусы, реже бактерий. Их присутсвие ведет к изменению жизнедеятельности клетки и развитию определенных признаков. Ложная цитоплазматическая наследственность (рак молочной железы у мышей с молоком матери, так как вирус этот находится и в семенном пузырьке самцов, переносивших их при спаривании самкам). 2.Свойство цитоплазмы яйцеклетки определяется ядром матери. 3. непосредственное влияние материнского организма на развитие зародыша также нередко ведет к известному сходству потомства с матерью, хотя оно и необусловлено с наследственностью. Питание эмбриона, недоразвитость маток, крупная самка мелкий самец и наобарот, влияние маток – рецепиент на продолжительность плодоношения; мул – от кобылицы и осла крупнее и более сходен к лошади чем лошак – гибрид ослихи и жеребца. Типичная внеядерная, или цитоплазматическая наследственность может быть вызвана с одной стороны, влиянием органоидов, находящихся в цитомлазме, а с другой – некоторыми особенностями собственно плазмы, то есть гиало плазмы. Влияние на наследственность органоидов плазмы. Митохондрия – у животных и растений всех видов. Пластиды – в клетках зеленых растений (в которых синтез – ся хлорофил) У бактерии цитоплазма содержит небольшие молекула ДНК – ЭПИСОМЫ. Тоже подвержены мутациям которые передаются в дочерние клетки. Влияние наследственности цитоплазмы. Асатуров на тутовом шёлкопряде путем замены ядра не обнаружил влияние цитоплазмы. У амёбы разных видов установлено влияние плазмы на размер ядер и особенности псевдоподии. Заключение: Действие генов может быть иногда ослаблено или полностью подавлено. VІ. В кариотипе заложена генетическая информация особи, изменения которые влекут за собой изменения признаков и функций организма данной особи или ее потомства. Поэтому очень важно знать особенности нормального строения хоромосом, чтобы при возможности суметь выявить изменения в кариотипе. Паталогии митоза. При делении саматических клеток могут возникнуть различные нарушения, связанные с поврождением хромосом, митотического аппарата, цитоплазмы. К числу этих нарушений относятся: 1) задержка митоза в профазе, 2) нарушение спирализации и дестирилизации хромосом, 3) раннее разделение хроматид, 4) фрагментация и пульверизация хросом, 5) задержка митоза в метафазе, К – митоз и др. Эти нарушения возникают под действием отдельных химических веществ, радиации, вирусных инфекций. Так при заражении свиней вирусом чумы наблюдали пульверизацию и фрагментацию (множественные разрывы) хромосом. Патологии мейоза. Основная паталогия мейоза – нерасхождение хромосом. Полиплоидия – триплоидия, тетраплоидия, пентаплоидия, гексоплоидия. Анеуплоидия – (+) трисомия, полисомия, (-) моносомия, нуллисомия. V. Оплодотворение Под оплодотворением понимается слияние ядер спермия и яйцеклетки. Зигота имеет двойной набор хромосом – тем самым оплодотворение обеспечивает сочетание у потомка признаков и свойств его родителей. В яйцеклетку проникает несколько или много спермиев, но оплодотворение является моноспермным, а сверхкомплекстные спермии выполняют трофическую функцию, влияя на жизни способность развивающегося организма. И.И.Соколовская, Е.К.Меркурьева, Д.И.Генина (1950 – 1953) – проникновение спермиев в клетки половых путей самок, что указывает на их трофическую функцию, и проливает свет на значение избытка этих гамет. Случайность Избирательность: 1. межвидовая гибридизация 2. внутри одного вида Виды наследственности и изменчивости Виды наследственности Виды изменчивости Виды наследственности - Это свойство живых существ передовать свои признаки и свойства потомству. Различают:
Ядерная наследственность-обусловлена генами локализированными в хромосомах. Или определяется большая часть признаков или свойств организма. Внеядерная наследственность-обусловлена наличием в цитоплазме клеток органелл, имеющих собственную ДНК, а слдовательно собственные гены. К таким органелам относятя митохондрии, пластиды высших растений и базальные тельца, расположенные в основании жгутиков и ресничек. Выделют: 1)истинную 2) ложную 3) переходную наследственность
Ложная наследственность у живых наблюдается при заражении их медленными вирусами. Важно подчеркнуть, что собственно болезнь как комплекс признаков –симптомов заболевания-резултать действия генов «хозяйна» в ответ на активность генов возбудиться. Кроме того, и сама предрасположенность к заболеванию зависить от генов «хозяйка» Таким образом ложная наследственность не случайна, так как в основе мррио истенная наследственность, наличие генов орорроо к действую возбудителя болезни. Ложная наследственность проявляется у некоторых червей зеленая окраска тела которых определяется их симбионтами –одноклеточными зелеными водорослями. Окраска желтка яиц кур обусловлена поступлением в него каротине растительного поролдл При этом следует еще раз отметить, что возможность накопления в организме Вв, поступипшего губке, зависит от характера действия собственных генов организма.
Приобретенное «Чужое», становится элементом цельстной гентической системы «Хозяйка» ІІ Виды изменчивости
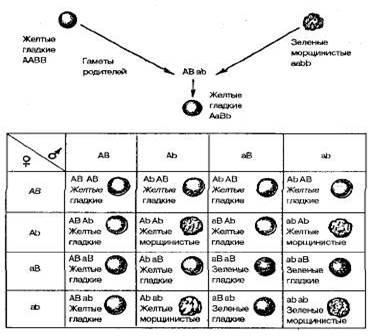
3. Закономерности наследования признаков при половом размножении -понятия ген, аллели-аллель-ные гены, аллелеморфы- множественный аллелизм, неаллельные гены, гомо- и гетерозиготе, генотипе и фенотипе, доминантности и рецессивности; -моногибридное скрещивание; -взаимодействие аллельных генов; -реципикорные и возвратные - анализирующие скрещивания; -ди- и полигибридные скрещиваниях, -взаимодействие неаллельных генов Дигибридное скрещивание Сущность дигибридного скрещивания. Организмы различаются по многим генам и, как следствие, по многим признакам. Чтобы одновременно проанализировать наследование нескольких признаков, необходимо изучить наследование каждой пары признаков в отдельности, не обращая внимания на другие пары, а затем сопоставить и объединить все наблюдения. Именно так и поступил Мендель. Скрещивание, при котором родительские формы отличаются по двум парам альтернативных признаков (по двум парам аллелей), называется дигибридным. Гибриды, гетерозиготные по двум генам, называют дигетерозиготными, а в случае отличия их по трем и многим генам —три- и полигетерозиготными соответственно. Результаты дигибридного и полигибридного скрещивания зависят от того, располагаются гены, определяющие рассмотренные признаки, в одной хромосоме или в разных. Независимое наследование (третий закон Менделя). Для дигибридного скрещивания Мендель использовал гомозиготные растения гороха, различающиеся одновременно по двум парам признаков. Одно из скрещиваемых растений имело желтые гладкие семена, другое — зеленые морщинистые (рис. 3.3).
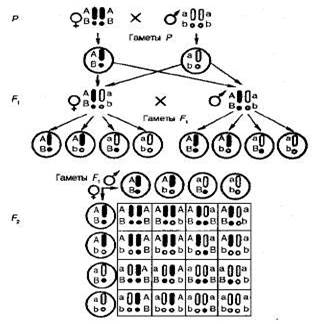
Рис 3.3. Дигибридное скрещивание растений гороха, различающихся по форме и окраске семян. Все гибриды первого поколения этого скрещивания имели желтые гладкие семена. Следовательно, доминирующими оказались желтая окраска семян над зеленой и гладкая форма над морщинистой. Обозначим аллели желтой окраски А, зеленой — а, гладкой формы— В, морщинистой— b. Гены, определяющие развитие разных пар признаков, называются неаллельпыми и обозначаются разными буквами латинского алфавита. Родительские растения в этом случае имеют генотипы АА ВВ и aabb, а генотип гибридов F1 —АаВb, т. е. является дигетерозиготным. Во втором поколении после самоопыления гибридов F1 в соответствии с законом расщепления вновь появились морщинистые и зеленые семена. При этом наблюдались следующие сочетания признаков: 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых семян. Это соотношение очень близко к соотношению 9: 3: 3: 1. Чтобы выяснить, как ведет себя каждая пара аллелей в потомстве дигетерозиготы, целесообразно провести раздельный учет каждой пары признаков — по форме и окраске семян. Из 556 семян Менделем получено 423 гладких и 133 морщинистых, а также 416 желтых и 140 зеленых. Таким образом, и в этом случае соотношение доминантных и рецессивных форм по каждой паре признаков свидетельствует о моногибридном расщеплении по фенотипу 3: 1. Отсюда следует, что дигибридное расщепление представляет собой два независимо идущих моногибридных расщепления, которые как бы накладываются друг на друга. Проведенные наблюдения свидетельствуют о том, что отдельные пары признаков ведут себя в наследовании независимо. В этом сущность третьего закона Менделя — закона независимого наследования признаков, или независимого комбинирования генов. Он формулируется так: каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга. Закон независимого комбинирования генов составляет основу комбинативной изменчивости (см. § 3.4), наблюдаемой при скрещивании у всех живых организмов. Отметим также, что в отличие от первого закона Менделя, который справедлив всегда, второй закон действителен только для генов, локализованных в разных парах гомологичных хромосом. Это обусловлено тем, что негомологичные хромосомы комбинируются в клетке независимо друг от друга, что было доказано не только при изучении характера наследования признаков, но и прямым цитологическим методом. Поведение хромосом при дигибридном скрещивании показано на рис. 3.4.
Цитологические основы дигибридного скрещивания. Как известно, в профазе I мейоза гомологичные хромосомы конъюги-руют, а в анафазе одна из гомологичных хромосом отходит к одному полюсу клетки, а другая — к другому. При расхождении к разным полюсам негомологичные хромосомы комбинируются свободно и независимо друг от друга. При оплодотворении в зиготе восстанавливается диплоидный набор хромосом и гомологичные хромосомы, оказавшиеся в процессе мейоза в разных половых клетках родителей, соединяются вновь. Предположим, что каждая хромосома содержит только один ген. Палочковидные хромосомы несут аллель A или а, сферические —В или b, т. е. эти две пары аллелей находятся в негомологичных хромосомах Гомозиготные родители (ААВВ и aabb) формируют только один тип гамет с доминантными (АВ) или с рецессивными (ab) аллелями. При слиянии таких гамет образуется единообразное первое поколение гибридов — гибрид дигетерозиготен (АаВb), но так как у него присутствуют гены А и B, то по фенотипу он сходен с одним из родителей. В тех случаях, когда необходимо указать, что те или иные гены находятся в гомологичных хромосомах, в генетических формулах зигот хромосомы принято изображать в виде двух черточек или одной с указанием обоих аллелей гена. Формула дигетерозиготы может быть записана так. Поскольку гаметы содержат только по одной из гомологичных хромосом и соответственно по одному аллелю каждого гена, то их формулы могут быть записаны так: и т. д. В дальнейшем у гибридных организмов ло причине случайности расхождения отцовских и материнских хромосом каждой пары в процессе мейоза ген А может попасть в одну гамету с геном В или с геном Ь. Точно так же ген а может оказаться в одной гамете с геном В или с геном b. Поэтому гибриды образуют четыре типа гамет: Образование всех четырех типов гамет равновероятно, т. е. все они образуются в равных количествах. Свободное сочетание таких гамет в процессах оплодотворения заканчивается образованием 16 типов зигот, а значит, и потомков. Они распадаются на четыре фенотипических класса: доминантные по обоим признакам — 9 частей, доминантные по первому и рецессивные по второму признаку — 3 части, рецессивные по первому и доминантные по второму — 3 части, рецессивные по обоим признакам — 1 часть. Генотипических классов 9: 1AABB, 2ААВb, 1AAbb, 1Aabb, 4AaBb, 2AaBB, 1aaBB, 2aaBb, 1aabb. Полигибридное скрещивание. Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании, т. е. когда родители различаются по аллелям трех и более генов, а в F1 образуются три- и по дигетерозиготы. Соотношение генотипических и фенотипических классов в F2 три- и полигибридных скрещиваний, а также число типов гамет (и число фенотипов) у гибридов F1 определяются простыми формулами: примоногибридном скрещивании число типов гамет равно 2, при дигибридном 4(22), а при полигибридном — 2n; число генотипов равно соответственно 3, 9(32) и 3n. Опираясь на независимость наследования разных пар аллелей, можно также любые сложные расщепления представить как произведение от соответствующего числа независимых моногиб-ридиых скрещивании. Общая формула определения фенотипических классов при полигибридном скрещивании имеет вид (3: 1)n, где п равно числу пар признаков, по которым идет расщепление. Для моногибрида эта формула соответственно имеет вид (3: 1); дигибрида — 9: 3: 3: 1 или(3: 1)2; тригибрида — (3: 1)3. Расщепление по генотипу имеет вид (1: 2; 1)n, где п — число расщепляющихся пар аллелей. Известно, что каждый организм гетерозиготен по многим генам. Если предположить, что человек, у которого отдельные пары хромосом содержат не одну, а сотни пар аллелей, гетерозиготен хотя бы по 20 генам, то число типов гамет у такой полигетерозиго-ты составит 220 = 1 048 576. Эта цифра дает определенное представление о потенциальных возможностях комбинативной изменчивости. Поэтому каждый человек обладает неповторимой индивидуальностью. На Земле нет двух людей, совершенно одинаковых по наследственности, за исключением однояйцевых близнецов. Таким образом, третий закон Менделя (закон независимого наследования признаков) еще раз демонстрирует дискретный характер генетического материала. Это проявляется в независимом комбинировании аллелей разных генов и в их независимом дей-ствии — фенотипическом выражении. Дискретность гена определяется тем, что он контролирует присутствие или отсутствие отдельной биохимической реакции, от которой зависит развитие или подавление определенного признака организма. Очевидно, если несколько генов определяют какое-либо одно свойство или один признак (форма гребня у кур, окраска глаз у дрозофилы, длина колоса у пшеницы и т. д.), они должны взаимодействовать между собой. Отсюда следует, что понятие «наследование признаков» употребляется, скорее всего, как образное выражение, поскольку в действительности наследуются не сами признаки, а гены. Признаки формируются в ходе индивидуального развития организма, обусловливаются генотипом и влиянием внешней среды. 4. Изучение изменчивости признаков методами вариационной статистики. -понятия о генеральной и выборочных совокупностях, -вариационный ряд и его графическое изоображение, -вычисление статистических показателей большой и малой выборок, -определение уровня вероятности статистических показателей и достоверности различии между группами животных, -доверительнее границы и правило трёх сигм, -критерий хи-квадрат, -коррелятивная связь между признаками.
5. Хромосомная теория наследственности -сцепленное наследование признаков, -кроссинговер, -хромосомная теория наследственности; Хромосомная теория наследственности Основоположник теории Томас Гент Морган, американский генетик, нобелевский лауреат, выдвинул гипотезу об ограничении законов Менделя. В экспериментах он использовал плодовую мушку-дрозо-филу, обладающую важными для генетических экспериментов качествами: неприхотливостью, плодовитостью, небольшим количеством хромосом (четыре пары), множеством четко выраженных альтернативных признаков. Морган и его ученики установили следующее: 1. Гены, расположенные в одной хромосоме, наследуются совместно или сцепленно. 2. Группы генов, расположенных в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и п+1 у гетерогаметных особей. 3. Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссин-говера возникают гаметы, хромосомы которых содержат новые комбинации генов. 4.Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей. При значении этой величины в 10 морганид можно утверждать, что частота перекреста хромосом в точках расположения данных генов равна 10% и что в 10% потомства будут выявлены новые генетические комбинации. 5. Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы. Эти выводы Моргана и его сотрудников получили название хромосомной теории наследственности. Важнейшими следствиями этой теории являются современные представления о гене как о функциональной единице наследственности, его делимости и способности к взаимодействию с другими генами. Пример сцепленного наследования: Vg — нормальные крылья дрозофилы; vg — зачаточные крылья; ВВ— серая окраска тела; bb — темная окраска тела. Запись в хромосомном выражении:
В данном случае правило единообразия гибридов первого поколения соблюдается. В соответствии со вторым и третьим законами Менделя следовало ожидать при последующем анализирующем скрещивании по 25% каждого из возможных фенотипов (серых, длиннокрылых мух, серых короткокрылых, черных длиннокрылых и черных коротко-крылых). Однако опыты Моргана не дали таких результатов. При скрещивании рецессивной по обоим признакам самки VgVgbb с гибридным самцом из F1 образовалось 50% серых мух с короткими крыльями и 50% мух с черным телом и длинными крыльями:
Если же скрещивают дигибридную самку с гомозиготным рецессивным самцом, то в образуется потомство: 41, 5% — серых с короткими крыльями, 41, 5% — черных с длинными крыльями, 8, 5% — серых с длинными крыльями, 8, 5% — черных с короткими крыльями. Данные результаты свидетельствуют о наличии сцепления генов и кроссинговере между ними. Так как в потомстве от второго скрещивания было получено 17% рекомбинант-ных особей, то расстояние между генами Vg и В равно 17%, или 17 морганидам. Наследование, сцепленное с полом Хромосомные наборы разных полов отличаются по строению половых хромосом. У-хромосома мужчин не содержит многих аллелей, имеющихся в Х-хромосоме. Признаки, определяемые генами половых хромосом, называют сцепленными с полом. Характер наследования зависит от распределения хромосом в мейозе. У гетерогаметных полов признаки, сцепленные с Х-хромосомой и не имеющие аллеля в У-хро-мосоме, проявляются даже в том случае, когда ген, определяющий развитие этих признаков, — рецессивен. У человека У-хромосома передается от отца к сыновьям, а Х-хромосо-ма — к дочерям. Вторую хромосому дети получают от матери. Это всегда Х-хромосома. Если мать несет патологический рецессивный ген в одной из Х-хромосом (например, ген дальтонизма или гемофилии), но при этом сама не больна, то она является носительницей. В случае передачи этого гена сыновьям они могут родиться с данным заболеванием, ибо в У-хромосоме нет аллеля, подавляющего патологический ген. Пол организма определяется в момент оплодотворения и зависит от хромосомного набора образовавшейся зиготы. У птиц гетерогаметными являются самки, а гомогаметными — самцы. У пчел половых хромосом вообще нет. Самцы гаплоидны. Самки пчел диплоидны. Пример наследования, сцепленного с полом
Основные положения хромосомной теории наследственности: • каждый ген имеет в хромосоме определенный локус (место); • гены в хромосоме расположены в определенной последовательности; • гены одной хромосомы сцеплены, поэтому наследуются преимущественно вместе; • частота кроссинговера между генами равна расстоянию между ними; • набор хромосом в клетках данного типа (кариотип) является характерной особенностью вида. 6. Генетика пола -наследование пола; теории определения пола; -нарушения в развитии признаков пола, обусловленные патологиями набора половых хромосом; -фримартинизм; -гермофродитизм и гинандроморфизм; -проблемы регулирования пола у животных и пути ее решения; -особенности наследования сцепленных с полом признаков; -ограниченные полом признаки и их наследование. Генетика пола Наследование пола. Теории определения пола Различают три пути, опеределяющих формирование особей того или иного пола. 1.Прогамное определение пола. Характеризируется тем что формирование особей того или иного полаопределяется еще в организме самки и зависит от размера яйцеклеток. При этом из мелких яйцеклеток (и цитоплазма ала) после оплодотворения развиваются самцы, а из крупных самки. Наблюдается у коловороток, тлей и морских червей. 2.Эпигамное определение пола наступает после оплодотворения, на последующих стадиях развития особи. Обнаружена у морского червя БОНЕЛЛИА, развитие личинки этого червя может происходить двояко: - если личинка свободно плавает в морской воде и позднее оседает на дно, то она становится самкой; - если же личинка прикрепляется к хоботу взрослой самки, то под влиянием гормонов, выделяемых этой самки, из нее формируется самец. 3.Сингамное определение пола характеризуется тем, что пол будущей особи, определяется при оплодотворении гамет, в результате соответствующего сочетания половых хромосом, т.е. при образовании зиготы. Характерна для млекопитающих, птиц, рыб, двукрылых насекомых, двудомных растений. Сингамное определение пола объясняется двумя теориями: А) хромосомной теорией определения пола Б) балансовой теорией определения пола
Наблюдения, проведенные на животных разных видов, показали, что среди новорожденных 50% составляют самцы, и 50 % самки. Р ХХ * ХУ ххху гаметы хххххуху ♀ самки ♂ самцы 50% 50% Наследование пола. Половые хромосомы и аутосомы. Генотипический пол особи обусловлен различиями в хромосомном аппарате мужских и женских особей. Начиная 1901г появились предположения и цитологические подтверждения. (гомогаметный пол, гетерогаметный пол) Рыбы: - самцы - млекопитающих, большая часть насекомых (и дрозофила), малюски, нематод. - самки – птиц, пресмыкающихся, бабочки некоторых видов (тутовый шелкопряд). Х и У хромосомы почти не отличаются от аутосомы: 1) в них накоплен гены признаков; 2) линейность расположения генов; 3) происходит кроссинговер(когда ХХ), поэтому составлена карта хромосом; 4) морфологическая и химическая структура. Аутосомы и половые хромосомы
В Y– W тех генов, которые находятся в Х – Z нет, большая часть их является генетический пустой то есть, состоит в основном генетический инертного материала. Половой хроматин или тельце Барра (1949) = ГЕНЕТИКА ПОЛА
Интерсекс- это однополая особь, у который придаточные половые железы, половые органы или вторичные половые признаки изменены в направлении противоположного пола. Такихинтерсексов иногда называют псевдогермофрадитами. Фримартины- это гормональные интерсексы генетический женского пола из разнополых двоен. Среди телок из разнополых двоен 92% фримартины. Истинный гермофродит- это особь, который обладает как женскими так и мужскими половыми железами и продуцирует как яйцеклетки, так испермии. Пригина: химеризм соматических клеток организма по набору половых храмосом- ХХ и ХУ. Гинануроморф – особь, часть тела которого является генетический мужским, а другая часть – женским.
Аномалии, затрагивающие морфологические и физиологические системы.
Балансовая теория определения пола. В 1919 гБриджес предложил БАЛАНСОВУЮ теорию определения пола, согласно которой у дрозофилы, у кузнечика и некоторых других насекомых пол особи определяется соотношением аутосом и Х хромосомой. При этом предполагается, что гены обуславливающие развитие мужской особи локолизированы в аутосомах, а гены – развития женской особи в Х хромосомах. Дрозофила – по табл.плакату. ХХУ – самка нормальная ХХУ – самец (мужск.) ХО – самец бесплодный ХО – самка (женск.) У хромосома не определяет пол, а плодовитость. Набор Аутосома Х хромосома Аутосома Х хромосома (Х: Аутосома) 6: 2 2: 2 (1, 0) или 1: 1 самка - ♀ 6: 1 2: 1 (0, 5) 2: 1 самец - ♂ 9: 2 3: 2 (0, 67) интерсекс Интерсекс (от лат. inter — между и sexus — пол)– это однополая особь, у которой в той или иной степени развиты одновременно признаки как одного, так и другого пола, то есть у которой придаточные половые железы, половые органы или вторичные половые признаки изменены в направлении противоположного пола. Истинный гермофродит – это особь которая обладает как женскими, так и мужскими половыми железами и продуцирует как яйцеклетки, так и спермы. Гинандоморф – часть или некоторые части его тела является генетический мужскими, а часть генетический женскими. Фримартины – это гормональные интерсексы генетический женского пола из разнополых.
|
Последнее изменение этой страницы: 2017-03-17; Просмотров: 376; Нарушение авторского права страницы
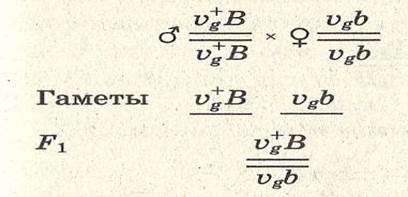
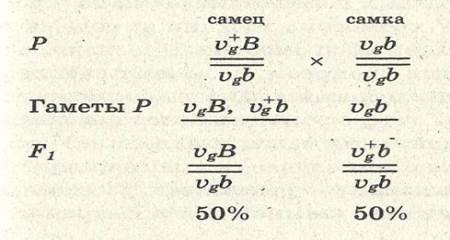

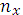 – 1.
– 1.