|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Перенос генетической информации путем конъюгации лучше всего изучен у энтеробактерий.
Конъюгация, как и другие виды рекомбинации, может осуществляться не только между бактериями одного и того же вида, но и между бактериями разных видов. В этих случаях рекомбинация называется межвидовой. ПЛАЗМИДЫ Плазмиды — это сравнительно небольшие внехромо-сомные молекулы ДНК бактериальной клетки. Они расположены в цитоплазме и имеют кольцевую структуру. В плазмидах содержится несколько генов, функционирующих независимо от генов, содержащихся в хромосомной ДНК. Типичным признаком плазмид служит их способность к самостоятельному воспроизведению (репликации). Они могут также переходить из одной клетки в другую и включать в себя новые гены из окружающей среды. К числу плазмид относятся: Профаги, вызывающие у лизогенной клетки ряд изменений, передающихся по наследству, например способность образовывать токсин (см. трансдукцию). F-фактор, находящийся в автономном состоянии и принимающий участие в процессе конъюгации (см. конъюгацию). R-фактор, придающий клетке устойчивость к лекарственным препаратам (впервые R-фактор был выделен из кишечной палочки, затем из шигелл). Исследования показали, что R-фактор может быть удален из клетки, что вообще характерно для плазмид. К-фактор обладает внутривидовой, межвидовой и даже межродовой трансмиссивностью, что может явиться причиной формирования трудно диагностируемых атипичных штаммов. Бактериоциногенные факторы (col-факторы), которые впервые были обнаружены в культуре кишечной палочки (E.coli), в связи с чем названы колицинами. В дальнейшем они были выявлены и у других бактерий: холерного вибриона — вибриоцины, стафилококков — стафилоцины и др. Соl-фактор — это маленькая автономная плазмида, которая детерминирует синтез белковых веществ, способных вызывать гибель бактерий собственного вида или близкородственного. Бактериоцины адсорбируются на поверхности чувствительных клеток и вызывают нарушения метаболизма, что приводит клетку к гибели. В естественных условиях только единичные клетки в популяции (1 на 1000) спонтанно продуцируют колицин. Однако при некоторых воздействиях на культуру (обработка бактерий УФ-лучами) количество колицинпродуцйрующих клеток увеличивается. ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ИЗМЕНЧИВОСТИ микроорганизмов Еще Пастер искусственным путем получил необратимые изменения у возбудителей бешенства, сибирской язвы и приготовил вакцины, предохраняющие от этих заболеваний. В дальнейшем исследования в области генетики и изменчивости микроорганизмов позволили получить большое число бактериальных и вирусных штаммов, используемых для получения вакцин. Результаты исследования генетики микроорганизмов с успехом были использованы для выяснения закономерностей наследственности высших организмов. Большое научное и практическое значение имеет также новый раздел генетики — генная инженерия. Методы генной инженерии позволяют изменять структуру генов и включать в хромосому бактерий гены других организмов, ответственных за синтез важных и нужных веществ. В результате микроорганизмы становятся продуцентами таких веществ, получение которых химическим путем представляет очень сложную, а иногда даже невозможную задачу. Этим путем в настоящее время получают такие медицинские препараты, как инсулин, интерферон и др. При использовании мутагенных факторов и селекции были получены мутанты-продуценты антибиотиков, которые в 100—1000 раз активнее исходных. 9. Генетика иммунитета -генетическая детерминированность иммунной реакции организма высших животных; -механизм синтеза моноспецифических антител и иммунная память; -наследуемость уровня иммунной реакции организма и возможности селекции животных по устойчивости к инфекциям. Иммунитет – это невосприимчивость организма к инфекционным агентам и генетически чужеродным веществам антигенной природы. Главная функция иммунитета – иммунологический надзор за внутренним постоянством (гомеостазом) организма. Следствием этой функции является распознавание, а потом блокирование, нейтрализация или уничтожение генетически чужеродных веществ (вирусов, бактерий, раковых клеток и т.д.). За сохранение генетически обусловленной биологической индивидуальности отвечает иммунная система организма – совокупность всех лимфоидных клеток (специфический фактор защиты ). К неспецифическим факторам защиты относят кожные и слизистые покровы. Иммунный ответ, или иммунологическая реактивность – форма реакций организма на чужеродные вещества (антигены). Главной функцией антител является их способность вступать в быструю реакцию с антигеном в виде реакции глютинации, преципитации, лизиса, нейтрализации.
10. Группы крови и биохимический полиморфизм. -понятие о группах крови; -наследуемость групп крови; -практическое применение групп крови в животноводстве; -полиморфные системы белков и их связь с продуктивностью животных; -методы определения групп крови и полиморфных систем белков. Группы крови были открыты в 1900 г. (у человека) и объяснены в 1924 г. А в 1936 году использован термин иммуногенетика. В пределах вида особи различаются по ряду охимических, генетически детерминируемых признаков, которые могут быть выявлены иммуногенетически в виде антигенов (генетически чужеродные вещества, при введении их в организм вызывают иммуногенетических реакций). Антитела – иммуноглобулины (белки), образующие в организме под воздействием антигенов, различия в групповой принадлежности крови определяются антигенами, расположенными на поверхности эритроцитов. Антигенные факторы иногда называют кровяными факторами, сумму всех групп крови одной особи – типом крови. После рождения группы крови у животных не меняется.. Генетические системы групп крови и антигены обозначают прописными и строчными буквами – А, В, С и т.д. Количество антигенов много, поэтому пишут со значками А, В, С, и с подстрочными индексами А1, А2 и т.д. У всех видов животных большинство аллелей генетических систем групп крови наследуется по типу кодоминирования, т.е. в гетерозиготе фенотипически проявляются оба гена. Все известные системы групп крови у с/х животных локализованы в аутосомах. Можно выделить три правила наследования групп крови: 1) каждая особь наследует по одному из двух аллелей от матери и отца по каждой системе групп крови; 2) Особь с антигенами, не обнаруженными хотя бы у одного из родителей, не может быть потомком данной пары родителей; 3) Гомозиготная особь по одному антигену, например F/F, не может быть потомком гомозиготной особи с противоположным антигеном, например V/V. В настоящее время у крупного рогатого Контроль достоверности происхождения животных возможен благодаря: 1) кодоминантному наследованию антигенных факторов; 2) их неизменности в течение онтогенеза; 3) огромному числу комбинации групп крови, которые в пределах вида не бывают одинаковыми у двух особей, за исключением монозиготных близнецов. Полиморфизм – это одновременное присутствие двух или более генетических форм одного вида. Термин генетический (биохимический) полиморфизм применяется тогда, когда локус хромосомы в популяции имеет два или более аллелей. Ген, представленный более, чем одним аллелем, называют полиморфным геном.
11. Генетическая инженерия. -генная инженерия: получение рекомбинантных молекул, внедрение рекомбинантных молекул, производство продуктов с помощью ГеноМодифицированных клеток и Организмов; -клеточная инженерия: получение моноклональных антител на основе гибридом; использование мезенхимальных и эмбриональ-ных стволовых клеток в лечении болезней и аномалии; выращивание культур клеток и клонирование низших организмов и высших животных.
Методы генной инженерии Генетическая инженерия - конструирование invitro функционально активных генетических структур (рекомбинантных ДНК), или иначе - создание искусственных генетических программ (Баев А. А.). По Э. С. Пирузян генетическая инженерия - система экспериментальных приемов, позволяющих конструировать лабораторным путем (в пробирке) искусственные генетические структуры в виде так называемых рекомбинантных или гибридных молекул ДНК. Генетическая инженерия - получение новых комбинаций генетического материала путем проводимых вне клетки манипуляций с молекулами нуклеиновых кислот и переноса созданных конструкций генов в живой организм, в результате которого достигается их включение и активность в этом организме и у его потомства. Речь идет о направленном, по заранее заданной программе конструировании молекулярных генетических систем вне организма с последующим введением их в живой организм. При этом рекомбинантные ДНК становятся составной частью генетического аппарата рецепиентного организма и сообщают ему новые уникальные генетические, биохимические, а затем и физиологические свойства. Цель прикладной генетической инженерии заключается в конструировании таких рекомбинантных молекул ДНК, которые при внедрении в генетический аппарат придавали бы организму свойства, полезные для человека. Например, получение «биологических реакторов» - микроорганизмов, растений и животных, продуцирующих фармакологически значимые для человека вещества, создание сортов растений и пород животных с определёнными ценными для человека признаками. Методы генной инженерии позволяют провести генетическую паспортизацию, диагностировать генетические заболевания, создавать ДНК-вакцины, проводить генотерапию различных заболеваний. Технология рекомбинантных ДНК использует следующие методы:
История генной инженерии Генная инженерия появилась благодаря работам многих исследователей в разных отраслях биохимии и молекулярной генетики. На протяжении многих лет главным классом макромолекул считали белки. Существовало даже предположение, что гены имеют белковую природу. Лишь в 1944 году Эйвери, Мак Леод и Мак Карти показали, что носителем наследственной информации является ДНК. С этого времени начинается интенсивное изучение нуклеиновых кислот. Спустя десятилетие, в 1953 году Дж. Уотсон и Ф. Крик создали двуспиральную модель ДНК. Именно этот год принято считать годом рождения молекулярной биологии. На рубеже 50 - 60-х годов были выяснены свойства генетического кода, а к концу 60-х годов его универсальность была подтверждена экспериментально. Шло интенсивное развитие молекулярной генетики, объектами которой стали E. coli, ее вирусы и плазмиды. Были разработаны методы выделения высокоочищенных препаратов неповрежденных молекул ДНК, плазмид и вирусов. ДНК вирусов и плазмид вводили в клетки в биологически активной форме, обеспечивая ее репликацию и экспрессию соответствующих генов. В 70-х годах был открыт ряд ферментов, катализирующих реакции превращения ДНК. Особая роль в развитии методов генной инженерии принадлежит рестриктазам и ДНК-лигазам. Историю развития генетической инженерии можно условно разделить на три этапа. Первый этап связан с доказательством принципиальной возможности получения рекомбинантных молекул ДНК invitro. Эти работы касаются получения гибридов между различнымиплазмидами. Была доказана возможность создания рекомбинантных молекул с использованием исходных молекул ДНК из различных видов и штаммов бактерий, их жизнеспособность, стабильность и функционирование. Второй этап связан с началом работ по получению рекомбинантных молекул ДНК между хромосомными генами прокариот и различными плазмидами, доказательством их стабильности и жизнеспособности. Третий этап - начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-рецепиента) генов эукариот, главным образом, животных. Формально датой рождения генетической инженерии следует считать 1972 год, когда в Стенфордском университете П. Берг, С. Коэн, Х. Бойер с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli. Читать дальше ► ферменты для манипуляций с ДНК и РНК Генетическая инженерия - потомок молекулярной генетики, но своим рождением обязана успехам генетической энзимологии и химии нуклеиновых кислот, так как инструментами молекулярного манипулирования являются ферменты. Если с клетками и клеточными органеллами мы подчас можем работать микроманипуляторами, то никакие, даже самые мелкие микрохирургические инструменты не помогут при работе с макромолекулами ДНК и РНК. Что же делать? В роли " скальпеля", " ножниц" и " ниток для сшивания" выступают ферменты. Только они могут найти определенные последовательности нуклеотидов, " разрезать" там молекулу или, наоборот, " заштопать" дырку в цепи ДНК. Эти ферменты издавна работают в клетке, выполняя работы по репликации (удвоению) ДНК при делении клетки, репарации повреждений (восстановлению целостности молекулы), в процессах считывания и переноса генетической информации из клетки в клетку или в пределах клетки. Задача генного инженера - подобрать фермент, который выполнил бы поставленные задачи, то есть смог бы работать с определенным участком нуклеиновой кислоты. Следует отметить, что ферменты, применяемые в генной инженерии, лишены видовой специфичности, поэтому экспериментатор может сочетать в единое целое фрагменты ДНК любого происхождения в избранной им последовательности. Это позволяет генной инженерии преодолевать установленные природой видовые барьеры и осуществлять межвидовое скрещивание. Ферменты, применяемые при конструировании рекомбинантных ДНК, можно разделить на несколько групп: - ферменты, с помощью которых получают фрагменты ДНК (рестриктазы); - ферменты, синтезирующие ДНК на матрице ДНК (полимеразы) или РНК (обратные транскриптазы); - ферменты, соединяющие фрагменты ДНК (лигазы); - ферменты, позволяющие осуществить изменение структуры концов фрагментов ДНК.
Рестриктазы Рестриктазы (рестрицирующиеэндонуклеазы, эндонуклеазы рестрикции) - это ферменты, узнающие и атакующие определенные последовательности нуклеотидов в молекуле ДНК (сайты рестрикции). Еще в 1953 году было обнаружено, что ДНК определенного штамма E. coli, введенная в клетки другого штамма (например, ДНК штамма В - в клетки штамма С) не проявляет, как правило, генетической активности, так как быстро расщепляется на мелкие фрагменты. В 1966 году было показано, что это явление связано со специфической модификацией хозяйской ДНК - она содержит несколько метилированных оснований, отсутствующих в немодифицированной ДНК, причем метилирование (добавление к основанию метильной группы) происходит уже после завершения репликации. Бактерия способна отличить свою собственную ДНК от любой вторгающейся «чужеродной» именно по типу ее модификации. За «метку» отвечают метилирующие ферменты модификации, так называемые ДНК-метилазы. Различие в модификации делает чужеродную ДНК чувствительной к действию рестрицирующих ферментов, которые узнают отсутствие метильных групп в соответствующих сайтах. Системы рестрикции и модификации широко распространены у бактерий; их существование играет важную роль в защите резидентной ДНК от загрязнения последовательностями чужеродного происхождения. Рестриктаза, которая расщепляла неметилированную ДНК была выделена в 1968 г. Мезельсоном и Юанем. Этот фермент был высокоспецифичен по отношению к определенной последовательности ДНК, но расщеплял молекулы неспецифически, в другом месте, на некотором удалении от участка узнавания. Вскоре, в 1970 г. Смит и Вилькокс выделили из Haemophilusinfluenzae первую рестриктазу, которая расщепляла строго определенную последовательность ДНК (Hind III). Поскольку разные бактерии по-разному метят свою ДНК, то и рестриктазы должны узнавать разные последовательности. И действительно, с тех пор выделены рестриктазы, узнающие более 150 сайтов рестрикции (мест расщепления ДНК).
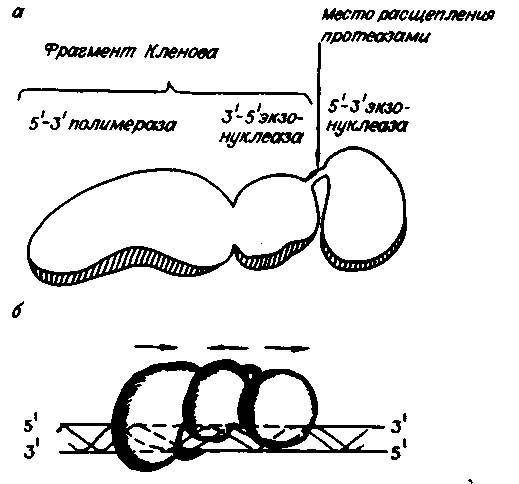
Полимеразы Впервые ДНК-полимераза была выделена Корнбергом с сотрудниками в 1958 году из E. coli. ДНК-полимераза I E. coli (Pol I) не связывается с молекулами двухцепочечной кольцевой ДНК. Однако, если такие молекулы денатурировать и получить одноцепочечные формы, то с последними полимераза связывается в количествах, пропорциональных длине этих участков — примерно одна молекула на 300 нуклеотидных остатков. Pol l связывается с одноцепочечными участками двойной спирали ДНК, в местах одноцепочечных разрывов с З'-гидроксилом и 5'-фосфатом, а также с концами двухцепочечных молекул ДНК. Фермент состоит из мономерной полипептидной цепи с молекулярной массой 103 кДа и имеет 3-х доменную структуру. Каждый домен обладает своей ферментативной активностью: 5’ - 3’ полимеразной, 3’ - 5’ экзонуклезной, 5’ - 3’ экзонуклеазной. 1. 5'— 3' полимеразная активность. Для реакции необходимо наличие одноцепочечной ДНК-матрицы и комплементарного участку этой цепи фрагмента — праймера (затравки) с З'-ОН концом. 2. 3'- 5' экзонуклеазная активность. Гидролизуетодноцепочечную или двухцепочечную ДНК с З'-ОН конца. 3'—5' нуклеаза расщепляет диэфирную связь только в неспаренных участках ДНК. Известно, что при полимеразной реакции с определенной частотой возможно включение в растущую цепь некомплементарного нуклеотида. Однако полимераза не может присоединять нуклеотид к неправильно спаренному концу, образовавшемуся при ее участии. На помощь приходит 3'—5' экзонуклеаза, убирающая ошибочный нуклеотид, на место которого затем присоединяется правильный нуклеотид-предшественник. 3'—5' экзонуклеолитическая активность проявляется в направлении, обратном синтезу ДНК (см. рис. 34). Таким образом, 3'—5' экзонуклеазная активность ДНК-полимеразы играет важную роль в точности полимеризации, направляемой матрицей. Эффективность, или число оборотов, данной экзонуклеазы в оптимальных условиях составляет 2% от числа оборотов субъединицы с полимеразной активностью. 3. 5'— 3' экзонуклеазная активность. Деградирует одну цепь двухцепочечной ДНК, начиная со свободного 5'-конца. В отличие от 3'—5' экзонуклеазы 5'—3' экзонуклеаза расщепляет диэфирную связь только в спаренных участках двухцепочечной молекулы ДНК. Более того, в то время как 3'—5' нуклеаза отщепляет одномоментно только один нуклеотид, 5'—3' нуклеаза может вырезать с 5'- конца олигонуклеотиды длиной до десяти остатков (около 20% продуктов гидролиза): Скорость нуклеазного отщепления увеличивается на порядок при одновременно протекающей реакции полимеризации. При этом увеличивается относительное количество олигонуклеотидов в продуктах гидролиза ДНК. Такое сочетание ферментативных активностей позволяет ДНК-полимеразе I E. coli играть активную роль в репарации повреждений ДНК invivo. N - концевой домен соединен с соседним петлей из аминокислотных остатков и легко отделяется с помощью протеолитических ферментов. Оставшаяся часть бифункциональна, так как состоит из полимеразы и 3’ - 5’ экзонуклезы. Она названа фрагментом Кленова (по фамилии одного из авторов, описавших ее). Фрагмент Кленова (Pol IK) обычно используют для достройки одноцепочечных 5'-концов на двухцепочечной ДНК, часто генерируемых рестриктазами, до тупых; для синтеза второй цепи на одноцепочечной ДНК, а также для гидролиза одноцепочечных З'-концов на двухцепочечных молекулах ДНК.
Обратная транскриптаза Обратная транскриптаза используется для транскрипции м-РНК в комплементарную цепь ДНК. При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза. Наиболее детально изучена ревертаза ретровирусов птиц. Каждый вирион содержит около 50 молекул этого фермента. Обратная транскриптаза состоит из двух субъединиц — a (65 кДа) и b (95 кДа), присутствующих в эквимолярном количестве. Обратная транскриптаза обладает, по крайней мере, тремя ферментативными активностями: 1) ДНК-полимеразной, использующей в качестве матрицы как РНК, так и ДНК; 2) активностью РНКазы Н, гидролизующей РНК в составе гибрида РНК - ДНК, но не одно- или двухцепочечную РНК; 3) ДНК-эндонуклеазной активностью. Первые две активности необходимы для синтеза вирусной ДНК, а эндонуклеаза, по-видимому, важна для интеграции вирусной ДНК в геном клетки-хозяина. Очищенная обратная транскриптаза синтезирует ДНК как на РНК-, так и на ДНК-матрицах. Чтобы начать синтез, ревертазе, как и другим полимеразам, необходим короткий двухцепочечный участок (праймер). Праймером может служить одноцепочечный сегмент как РНК, так и ДНК, которые в процессе реакции оказываются ковалентно связанными с новосинтезированной цепью ДНК.
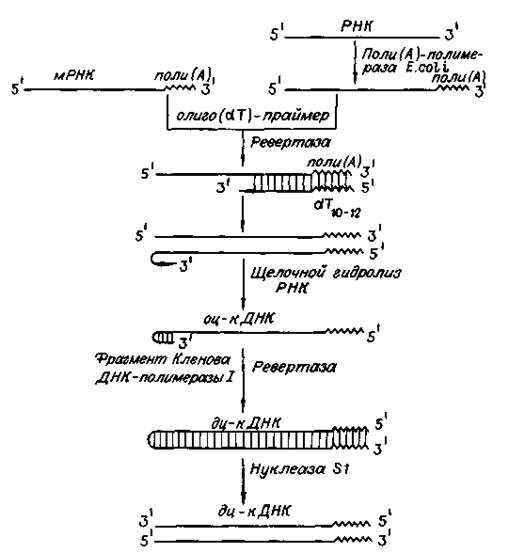
Рис. 35. Схема синтеза двухцепочечных ДНК-копий молекул РНК Обратную транскриптазу преимущественно используют для транскрипции матричной РНК в комплементарную ДНК (кДНК). Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А)-содержащих мРНК используют олигo (dT), а для молекул РНК, не имеющих З'-поли (А) концов, — химически синтезированные олигонуклеотиды, комплементарные З'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечныхкДНКсамокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I E. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающейодноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимеразы I E. coli. Полученную двухцепочечнуюкДНК можно затем встраивать в клонирующие векторы, размножать в составе гибридных молекул ДНК и использовать для дальнейших исследований. Лигазы В 1961 г. Мезельсон и Вейгл на примере фага l показали, что рекомбинация включает разрыв и последующее воссоединение молекул ДНК. Это положило начало поискам фермента, участвующего в сшивании фрагментов ДНК. В 1967 году такой фермент был найден и получил название ДНК-лигаза. Он катализирует синтез фосфодиэфирной связи в 2-х цепочечной молекуле нуклеиновой кислоты. Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя связь между остатками сахаров. ДНК-лигазы абсолютно необходимы в процессах репарации ДНК, в процессах репликации - при удвоении цепи ДНК. В генной инженерии используются 2 типа ДНК-лигаз, отличающихся по потребностям в кофакторах и способу действия. ДНК-лигаза E. coli в качестве кофактора использует дифосфопиридиннуклеотид, а лигаза фага Т4 - АТФ в присутствии Mg2+. Лигаза фага Т4 более универсальна, так как помимо лигирования липких концов способна катализировать реакцию воссоединения двухцепочечных фрагментов ДНК с тупыми концами. Она используется чаще.
ДНК-лигаза, фото с сайта ru.wikipedia.org |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 146; Нарушение авторского права страницы
 © 1995-2010 Наталья Кузьмина
© 1995-2010 Наталья Кузьмина