
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Цепь клеточного окисления и места фосфорилирования АДФ
Митохондрии, их строение и функции
Совокупность реакций дыхательной цепи и процесс окислительного фосфорилирования происходят в митохондриях — субклеточных органеллах специфического строения. Количество их в клетке колеблется от нескольких сотен до нескольких десятков тысяч. Размеры и форма митохондрий различны в зависимости от типа клеток. Они бывают цилиндрической, сферической или нитевидной формы, а также пластинчатой и звёздчатой форм. Однако митохондрии, выделенные из различных источников, имеют много общего. Оболочка этих органелл состоит из двух мембран — наружной и внутренней. Наружная мембрана гладкая, она как бы обтягивает и формирует митохондрию. Внутренняя мембрана образует перпендикулярные к поверхности мембраны многочисленные складки, или выпячивания, называемые кристами, которые вдаются в тело митохондрии довольно далеко, достигая даже противоположной стенки (рис. 42). Благодаря наличию крист внутренняя мембрана имеет очень большую протяженность. Внутреннее пространство митохондрий заполнено жидкостью, которая называется матриксом. Мембраны состоят на 1/3 из липидов и на 2/3— из белков, главным образом структурных. Около 25 % общего белка внутренней мембраны составляют белки ферментов, которые осуществляют перенос электронов и процессы форилирования. Этими ферментами являются НАД-зависимые дегидрогеназы, флавопротеиды, цитохромы b, с, а и а3.
В последнее время установлено, что ферменты митохондрии сгруппированы в компактные агрегаты с молекулярной массой 1 млн., которые называют дыхательными ансамблями (рис. 43). Дыхательные ансамбли вмонтированы поперек внутренней мембраны митохондрий и равномерно распределены в ней. Количество таких ансамблей в каждой митохондрии составляет 5000—10000. Следует отметить, что функциональная активность этих ансамблей проявляется только в соединении их с мембраной митохондрий. Отделение их от мембраны приводит к потере способности к сопряженному фосфорилированию хотя они и способны переносить электроны. Без мембраны отдельные ферменты дыхательной цепи становятся разобщенными, располагаются хаотично и бессистемно, в результате чего теряют способность утилизировать выделяемую при окислении энергию. В связи с этим считают, что функции митохондрий абсолютно уникальны. Поскольку именно в них происходит в основном синтез макроэргических соединений, митохондрии называют еще «силовыми станциями» клетки. Не следует думать, что любое окисление субстратов в живых системах сопровождаются фосфорилированием. Передача атомов водорода и электронов ферментами дыхательной цепи и связанное с этим фосфорилирование АДФ осуществляется в середине митохондрии, на ее внутренней мембране, где отдельные ферменты цепи переноса водорода и электронов соединены в строго определенном порядке, образуя своеобразный комплекс. Именно такое структурное сочетание ферментов с мембраной обеспечивает использование энергии окисления. Процесс передачи атомов водорода и электронов происходит и на поверхности митохондрий, и в гиалоплазме с участием тех же ферментов, что и окисление, сопряженное с фосфорилированием. Однако вследствиe того, что эти ферменты не связаны митохондриальной мембраной, они размещены хаотично и поэтому окисление здесь не сопровождается активацией неорганического фосфата. В этом случае энергия окисления превращается в теплоту итакой процесс называется свободным окислением. В настоящее время установлено, что процессы окислительного фосфорилирования и свободного окисления в клетках и в организме взаимосвязаны. При определенных условиях окислительное фосфорилирование может переключаться на свободное окисление. Такое переключение можно наблюдать при охлаждении организма, когда снижение температуры приводит к разобщению ферментов окислительного фосфорилирования и оно переходит в свободное окисление для усиления продукции теплоты. В заключение нужно сказать, что сопряженное окислительное фосфорилирование является основным, но не единственным способом синтеза АТФ в органах и тканях животного организма. Небольшое количество АТФ образуется путем так называемого субстратного фосфорилирования. В этом случае активирование неорганического фосфата происходит с участием самого субстрата. Поэтому такое фосфорилирование часто называют фосфорилированием на уровне субстрата, чтобы отличить его от реакций фосфорилирования, протекающих с участием дыхательной цепи. При субстратном фосфорилировании между субстратом и неорганическим фосфатом сначала образуется макроэргическая связь за счет окисления субстрата. Энергия его окисления передается остатку фосфорной кислоты. Активированный таким образом неорганический фосфат передается на АДФ с образованием АТФ: Субстрат (восстановленный) + Н3РО4 Субстрат (окисленный) ~ РО3Н2 + АДФ → АТФ + Окисленный субстрат. Субстратное фосфорилирование— очень древний способ биосинтеза АТФ. Он появился у организмов, когда на Земле не было кислорода, т.е. в анаэробных условиях. Однако клетки животного организма сохранили способность синтезировать АТФ путем субстратного фосфорилирования и осуществляют его при дефиците кислорода втканях. С примерами субстратного фосфорилирования мы ознакомимся в следующей главе. Глава 23. ОБМЕН УГЛЕВОДОВ Переваривание углеводов Углеводы занимают очень важное место в питании человека и животных. Они составляют большую часть (около 60-70 %) пищевого рациона человека и являются основным источником энергии, необходимой организму для осуществления разнообразных функций. Поэтому количество углеводов, как основного энергетического материала, должно увеличиваться в рационе человека по мере усиления физической нагрузки. Источником углеводов для человека являются продукты растительного происхождения — мука, различные крупы, картофель. Основным углеводом этих продуктов является крахмал и в незначительном количестве — клетчатка. С фруктами, ягодами и пищевыми сахарами человек получает преимущественно сахарозу и фруктозу. Однако не все углеводы усваиваются организмом человека в одинаковой степени. Клетчатка, например, не переваривается пищеварительными соками. Она лишь частично расщепляется при помощи бактерий в толстом отделе кишок до дисахарида целлобиозы и моносахарида глюкозы. Здесь же микроорганизмы используют клетчатку для биосинтеза витаминов группы К. Клетчатка необходима в качестве стимулятора секреторной имоторной функций пищевого канала и для формирования кала, поэтому она является важной составной частью пищевого рациона человека. Однако чрезмерное поступление клетчатки в организм вызывает усиление перистальтики кишок и пища проходит через пищевой канал очень быстро. В результате этого пищевые продукты не полностью перевариваются и всасываются, значительная часть их попадает в кал и удаляется из организма неиспользованной. Наряду с этим при повышенном количестве клетчатки в пищевом рационе в толстом отделе кишок усиливаются процессы брожения, происходит скопление газов, что может привести к серьезным осложнениям у больных людей, а также у тех, кто мало двигается. Такие полисахариды, как крахмал и гликоген, поступающие с пищей, предварительно подвергаются расщеплению в пищевом канале под действием определенных ферментов. В результате расщепления образуются моносахариды, которые затем легко ассимилируются тканями организма. Процесс переваривания углеводов (крахмала и дисахаридов) начинается уже в ротовой полости под действием двух ферментов, содержащихся в составе слюны, — амилазы и α -глюкозидазы (мальтазы). Амилаза расщепляет крахмал, α -глюкозидаза — мальтозу. Амилаза слюны является более активной по сравнению с мальтазой, поэтому в ротовой полости из крахмала образуются преимущественно большие фрагменты его молекул — декстрины, имеющие сладкий вкус, и лишь незначительное количество мальтозы и глюкозы. Поскольку в ротовой полости пища долго не задерживается, дальнейшее расщепление углеводов продолжается в следующих отделах пищевого канала. В желудке нет ферментов, расщепляющих углеводы. К тому же ферменты слюны прекращают свое действие в желудке, где реакция среды кислая, а оптимум действия этих ферментов лежит в нейтральной или слабокислой среде. В двенадцатиперстной кишке и в полости тонкой кишки под действием ряда ферментов поджелудочной железы и кишечного сока происходит окончательное расщепление углеводов до моносахаридов. Так, α -амилаза расщепляет крахмал и декстрины до мальтозы, α -глюкозидаза расщепляет мальтозу на две молекулы глюкозы. Под действием β -галактозидазы (лактазы) кишечного сока лактоза расщепляется на глюкозу и галактозу, β -фруктофуранозидаза (сахараза) расщепляет сахарозу на глюкозу и фруктозу. Образовавшиеся моносахариды, хорошо растворимые в воде, — глюкоза, фруктоза, галактоза— являются теми простыми сахарами, которые всасываются стенкой кишок и через капилляры кишечных ворсинок попадают в кровяное русло. Во время всасывания часть моносахаридов (фруктоза, галактоза и манноза) превращаются в глюкозу. Таким образом из кишок в кровь попадает преимущественно глюкоза и лишь в незначительных количествах — фруктоза, галактоза и манноза. С током крови через воротную вену они попадают в печень и другие органы и ткани. Часть глюкозы задерживается в печени и используется главным образом для биосинтеза гликогена и других сложных углеводов, часть превращается в жиры и окисляется в тканях. При нормальных условиях на синтез гликогена используется 3—5 % глюкозы, 30—35 % ее превращается в жиры, а основная масса — 60—70 % — окисляется до СО2 и Н2О с выделением энергии. Небольшое количество глюкозы является обязательной составной частью крови. В норме это составляет до 120 мг %. Этот уровень поддерживается несмотря на постоянное потребление глюкозы тканями и периодическое поступление ее из кишок после приема пищи. Главная роль в поддержании постоянной концентрации глюкозы в крови принадлежит печени. При недостаточном количестве глюкозы в крови под влиянием нервных импульсов, идущих из центральной нервной системы, часть гликогена печени расщепляется, повышая тем самым уровень глюкозы в крови. Этот процесс называют «мобилизацией» гликогена. Регуляторную роль в поддержании постоянного уровня глюкозы в крови центральная нервная система осуществляет не только путем прямого воздействия на печень, но и через эндокринные железы, среди которых наибольшее значение имеют надпочечники и поджелудочная железа. В мозговом веществе надпочечников образуется адреналин, повышенное содержание которого в крови стимулирует расщепление гликогена в печени. Поджелудочная железа вырабатывает гормон инсулин. Повышенное выделение этого гормона в кровь сопровождается снижением глюкозы в крови. Таким образом, инсулин действует противоположно адреналину — он задерживает распад гликогена в печени и способствует отложению его в мышцах. Кроме инсулина в поджелудочной железе вырабатывается еще один гормон — глюкагон, действие которого противоположно инсулину. Существенное влияние на уровень глюкозы в крови оказывает и ряд других гормонов: соматотропин, кортикостерон, тироксин. Все эти гормоны вызывают повышение содержания глюкозы в крови, в связи с чем их называют диабетогенными (от названия заболевания «диабет», сопровождаемого повышенным содержанием сахара в крови в результате нарушения функции поджелудочной железы, продуцирующей инсулин). Концентрация глюкозы в крови может увеличиваться до 160—180 мг % при употреблении с пищей большого количества углеводов. Это явление носит название алиментарной, или пищевой, гипергликемии, которая сопровождается выделением глюкозы с мочой, т.е. глюкозурией. Из вышесказанного следует, что поступление глюкозы в кровь происходит в основном двумя путями — расщеплением гликогена в печени и всасыванием простых сахаров из кишок после приема пищи. Образование глюкозы из гликогена происходит не прямым его расщеплением, а при помощи ферментов путем присоединения к нему фосфорной кислоты. При этом от гликогена последовательно отщепляются остатки глюкозы в виде фосфорных эфиров, которые затем расщепляются на глюкозу и фосфорную кислоту: (С6Н10О5)n (Гликоген)+ nН3РО4 Глюкозо-1-фосфат Небольшое количество глюкозы образуется также путем гидролитического расщепления гликогена, однако этот путь имеет второстепенное значение. |
Последнее изменение этой страницы: 2017-03-14; Просмотров: 593; Нарушение авторского права страницы

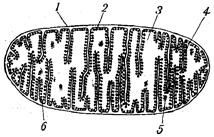 Рис. 42. Строение митохондрии: 1—наружная мембрана; 2— внутренняя мембрана; 3 — матрикс; 4 — кристы; 5 — внутримитохондриальные тельца: 6 — частицы внутренней мембраны (8, 0—9, 0 нм).
Рис. 42. Строение митохондрии: 1—наружная мембрана; 2— внутренняя мембрана; 3 — матрикс; 4 — кристы; 5 — внутримитохондриальные тельца: 6 — частицы внутренней мембраны (8, 0—9, 0 нм). Рис. 43. Схематическое изображение строения поверхности внутренней мембраны митохондрий: 1 - молекулы мембранных белков: НАД, ФАД, цитохромы b, с, а, а3, - ансамбль электронов; Х, У, Z - молекулы ферментов, ответственных за синтез АТФ.
Рис. 43. Схематическое изображение строения поверхности внутренней мембраны митохондрий: 1 - молекулы мембранных белков: НАД, ФАД, цитохромы b, с, а, а3, - ансамбль электронов; Х, У, Z - молекулы ферментов, ответственных за синтез АТФ. Субстрат(окисленный) ~ РО3Н2;
Субстрат(окисленный) ~ РО3Н2;  (C6H10O5)n-1 + Глюкозо-1-фосфат;
(C6H10O5)n-1 + Глюкозо-1-фосфат;  Глюкозо-6-фосфат
Глюкозо-6-фосфат  Глюкоза + H3PO4.
Глюкоза + H3PO4.