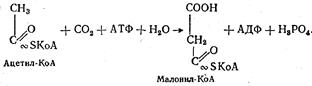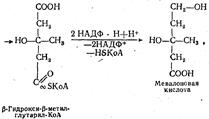|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Внутриклеточное окисление продуктов гидролитического расщепления жиров
Глицерин и высшие жирные кислоты, образовавшиеся в процессе пищеварения жиров, а также в результате расщепления триглицери-дов в жировых депо при участии тканевых липаз, подвергаются даль-нейшим превращениям. В клетках различных органов и тканей они окисляются до конечных продуктов (СО2 и Н2О) или в процессе окисления используются для биосинтеза других соединений. Окисление глицерина. Процесс окисления глицерина начинается фосфорилирования его под действием фермента глицеролкиназы. Донатором фосфорной кислоты в этой реакции выступает АТФ. В ре-зультате образуется глицеринфосфорная кислота:
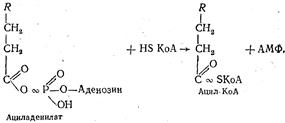
Наиболее интенсивно фосфорилирование глицерина происходит В клетках печени. Образовавшаяся глицеринфосфорная кислота при помощи глицеринфосфатдегидрогеназы превращается в диоксиацетон-фосфат, который изомеризуется в глицеральдегид-3-фосфат. Послед-ний подвергается окислению до фосфоглицериновой кислоты подобно тому, как это происходит в процессе гликолиза. Наряду с этим npoме-жуточные продукты окисления глицерина могут использоваться opгaнизмом для биосинтеза жиров, фосфоглицеридов и углеводов. Окисление высших жирных кислот. Все жирные кислоты перед окислением подвергаются активации. Этот процесс также происходит главным образом в печени и осуществляется с участием АТФ и коэнзима А. Схематически этот процесс можно представить следующим образом: сначала жирная кислота взаимодействует с АТФ с сбразозованием ациладенилата который, взаимодействуя с коэнзимом А, образует ацил-КоА— активную форму жирной кислоты: Ацил-КоА в дальнейшем вступает на путь окисления, который интенсивно протекает в митохондриях. Весь процесс окисления ацил-КоА заключается в постоянном дегидрировании его с отщеплением от ацил-КоА жирной кислоты двууглеродных фрагментов в виде ацетил-КоА (активной формы уксусной кислоты). Поскольку весь процесс дегидрирования сопровождается отщеплением от жирной кислоты двууглеродных фрагментов в β -положении, он носит название β -окисления. Дегидрирование ацил-КоА происходит под действием специфических дегидрогеназ с участием коферментов ФАД и НАД+: Кетоформа ацилкоэнзима А при помощи еще одной молекулы —КоА ферментативным путем расщепляется на две молекулы — ацетил-КоА и ацил-КоА: Укоротившись на два углеродных атома, молекула ацил-КоА снова подвергается двукратному дегидрированию с отщеплением новой молекулы ацетил-КоА. В конце концов вся молекула высшей жирной кислоты расщепляется до молекул ацетил-КоА. Если жирная кислота имела четное число атомов углерода, то в конце образуется целое число молекул ацетил-КоА. Так, молекула стеариновой кислоты (С17Н35СООН) образует 9 молекул, пальмитиновая (C15H31COOH) — 8 молекул ацетил-КоА. Образовавшиеся молекулы ацетил-КоА «сгорают» в цикле Кребса уже известным нам путем. При расщеплении высших жирных кислот до молекул ацетил-КоА высвобождается около 30 % энергии, остальные 70 % выделяются при окислении ацетил-КоА до СО2 и Н2О в цикле Кребса. Рассмотрим в качестве примера энергетический баланс окисления одной молекулы пальмитиновой кислоты, которая образует 8 молекул ацетил-КоА. Такое количество ацетил-КоА образуется в результате семи последовательных отщеплений, каждое из которых дважды сопровождается дегидрированием: один раз при участии ФАД, второй — при участии НАД. За счет каждой образовавшейся восстановленной формы ФАД•Н2 и НАД•Н + Н+ в дыхательной цепи образуется 5 молекул АТФ (ФАД•Н2 — 2 молекулы и НАД•Н + Н+ — 3). Всего же это составляет 35 молекул, так как отщепление ацетил-КоА произошло 7 раз (это почти столько же, сколько образуется при окислении молекулы глюкозы до СО2 и Н2О). За счет окисления каждой молекулы ацетил-КоА в цикле Кребса образуется еще 12 молекул АТФ, а в результате окисления 8 молекул — 96 молекул АТФ. Таким образом, окисление одной молекулы пальмитиновой кислоты сопровождается образованием 131 молекулы АТФ (35 + 96). Учитывая одну молекулу АТФ, использованную для активации пальмитиновой кислоты, «чистая прибыль» АТФ составит 130 молекул, в которых аккумулировано 4353—5442 кДж энергии. Поскольку в составе триглицерида содержится три остатка высшей жирной кислоты, а окисление глицерина также сопровождается образованием АТФ, общий итог окисления одной молекулы триглицерида будет еще большим. Биосинтез триглицеридов Синтез нейтральных жиров состоит из трех процессов: образования высшей жирной кислоты, образования глицерина и соединения этих веществ в молекулу триглицерида. Синтез высших жирных кислот. Местом образования высших жирных кислот является цитоплазма. В процессе участвуют активная форма уксусной кислоты, т.е. ацетил-КоА, и углекислый газ. На первом этапе биосинтеза при взаимодействии ацетил-КоА и СО2 образуется промежуточное соединение — малонил-КоА. Образование этого соединения происходит при участии витамина Н с использованием энергии АТФ:
В процессе биосинтеза высшей жирной кислоты малснил-КоА расщепляется на ацетил-КоА и СО2, а отдельные молекулы ацетил-КоА соединяются между собой в длинную цепь. Следовательно, малонил-КоА не входит в состав цепи высшей жирной кислоты, а служит лишь промежуточной формой, которая обеспечивает образование высшей жирной кислоты из отдельных молекул ацетил-КоА. Весь процесс- соединения молекул ацетил-КоА осуществляется с помощью фермента синтетазы жирных кислот, содержащей две сульфгидрильные группы: центральную и периферическую. На центральной тиоловой группе протекает реакция конденсации между малонил-КоА и ацетил-КоА с выделением СО2 и восстановление образовавшегося продукта, а периферическая группа служит для удержания образовавшейся цепи:
В результате соединения двух молекул ацетил-КоА и двух восстановительных реакций образовался фрагмент высшей жирной кислоты, состоящий из четырех атомов углерода. На следующем этапе образовавшийся фрагмент переносится с центральной тиоловой группы фермента на периферическую:
При этом центральная тиоловая группа освобождается и снова вступает в реакцию с новой молекулой малонил-КоА. Затем в такой же последовательности протекают реакция конденсации между образовавшейся цепью и молекулой малонил-КоА с выделением СО2 и реакции восстановления вновь образовавшегося продукта. Таким образом, при многоразовом повторении этих реакций углеводородная цепь все больше удлиняется, пока не образуется цепь из 16—18 атомов углерода. После этого синтезированная высшая жирная кислота взаимодействует с молекулой коэнзима А, образуя активную форму в виде ацил-КоА и освобождая при этом синтетазу жирной кислоты:
Биосинтез триглицеридов. Образование жира происходит в результате взаимодействия молекулы глицерина с тремя молекулами высших жирных кислот. Основным источником глицерина в организме служит промежуточный продукт окисления углеводов — диокси-ацетонмонофосфат, который путем восстановления превращается сначала в глицеринфосфорную кислоту, а затем в свободный глицерин. Субстратом в биосинтезе жирных кислот, как мы только что увидели, служит активная форма уксусной кислоты — ацетил-КоА. Как показали исследования, глицерин вступает в реакцию в виде глицеринфосфата, а жирные кислоты — в виде своей активной формы — ацил-КоА. На первом этапе биосинтеза триглицеридов происходит образование фосфатидной кислоты — общего промежуточного продукта в биосинтезе жиров и фосфатидов:
Далее фосфатидная кислота ферментативным путем расщепляется) на фосфорную кислоту и α -, β -диглицерид, который также является общим промежуточным продуктом в биосинтезе жиров и фосфатидов:
Образовавшийся α, β -диглицерид взаимодействует с третьей молекулой ацилкоэнзима А, образуя молекулу триглицерида:
Синтезированный таким путем специфический для организма жир откладывается в жировых депо. Биосинтез лецитина. Лецитин, как известно, принадлежит к группе фосфатидов. Для его биосинтеза необходимы следующие вещества: глицерин, высшие жирные кислоты, фосфорная кислота и азотистое основание холин. По своему строению лецитин до некоторой степени напоминает триглицериды. Поэтому многие этапы в биосинтезе лецитина аналогичны этапам биосинтеза нейтральных жиров. Как уже было сказано, общим промежуточным веществом в биосинтезе фосфатидов и триглицеридов является α, β -диглицерид. Как раз к этому промежуточному соединению и присоединяются фосфорная кислота и холин. Происходит это следующим образом. Вначале холин фосфорилируется при помощи АТФ с образованием фосфохолина:
который далее взаимодействует с цитидинтрифосфорной кислотой (ЦТФ) и превращается в цитидиндифосфохолин:
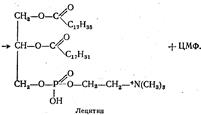
При взаимодействии цитидинфосфохолина с α, β -диглицеридом образуется лецитин:
Биосинтез лецитина очень активно протекает в стенке тонкой кишки и печени. Биосинтез холестерина. В организме человека холестерин синтезируется во всех органах и тканях. Механизм образования его очень сложен и долгое время оставался неясным. Благодаря использованию метода меченых атомов удалось установить все этапы его синтеза. Исходным веществом для синтеза холестерина служит активная форма уксусной кислоты — ацетил-КоА. Отдельные этапы его биосинтеза можно представить схематически следующим образом: 2 молекулы ацетил-КоА конденсируются с образованием ацетоацетил-КоА. К этой молекуле присоединяется третья молекула ацетил-КоА и образуется сложное соединение β -гидрокси-β -метилглутарил-коэнзим А. Из этого соединения образуется мевалоновая кислота. Образование мевалоновой кислоты схематически можно показать так:
Мевалоновая кислота является одним из важнейших промежуточных продуктов в биосинтезе холестерина. Через ряд промежуточных реакций она превращается в более сложное соединение. С помощью ЛТФ мевалоновая кислота превращается в свою активную форму — пирофосфат мевалоновой кислоты, который, дегидрируясь и декар-боксилируясь, образует изопентилпирофосфат. После изомеризации последнего образуется диметилаллилпирофосфат, который способен уже участвовать в реакциях конденсации и синтеза циклопентанпер-гидрофенантренового кольца холестерина. |
Последнее изменение этой страницы: 2017-03-14; Просмотров: 2474; Нарушение авторского права страницы