|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Аэробное окисление углеводов
Дыхание. Аэробное окисление углеводов происходит в присутствии кислорода воздуха, в связи с чем его часто называют дыханием. В отличие от гликолиза (гликогенолиза), где конечным акцептором атомов водорода и электронов служит провиноградная кислота, при дыхании роль такого акцептора выполняет кислород. В первом случае в качестве конечного продукта образуется молочная кислота, в которой суммарная степень окисления углерода осталась такой же, как и у глюкозы, во втором случае образуется углекислый газ — значительно более простое соединение, у которого единственный атом углерода полностью окислен. Вместе с тем дыхание и гликолиз имеют много общих звеньев. Дыхание, так же как и гликолиз, сопровождается образованием фосфорных эфиров глюкозы и фруктозы, фосфотриоз—диоксиацетонфосфата и глицеральдегид-3-фосфата, а также таких промежуточных продуктов, как 1, 3-дифосфоглицериновая кислота, 3-фосфоглицерат, фосфоенолпирувати пировиноградная кислота. Многие реакции гликолиза и дыхания катализируются одними и теми же ферментами. Другими словами, при дыхании превращение глюкозы до молочной кислоты проходит все те этапы, что и при гликолизе. Однако при этом атомы водорода, отщепленные от глицеральдегид-3-фосфата, не восстанавливают пировиноградную кислоту, а передаются на кислород, пройдя через сложную систему ферментов дыхательной цепи. Молочная кислота, образующаяся в процессе гликолиза, как уже говорилось, содержит еще довольно значительный запас (примерно 93 %) потенциальной энергии. Однако несмотря на это, первые живые организмы, извлекавшие энергию в анаэробных условиях, выделяли ее в окружающую среду. С появлением в атмосфере Земли кислорода живые организмы выработали новые, более совершенные механизмы окисления, в результате которых количество высвобождающейся энергии оказалось намного больше, чем при гликолизе, поскольку конечным продуктом дыхания является СО2, атом углерода которого полностью окислен. Наряду с этим природа создала новые механизмы доокисления конечного продукта гликолиза, который выводился в окружающую среду. Иными словами, она как бы создала надстройку над гликолизом для окисления его конечного продукта в аэробных условиях, сохранив прежними многие его этапы. При дыхании не образуется молочная кислота. Поэтому пировиноградная кислота является тем общим субстратом, или центральным звеном, где заканчивается гликолиз и начинается дыхание (или же расходятся пути гликолиза и дыхания — анаэробного и аэробного окисления глюкозы). Сохранив прежние этапы гликолиза, клетки организма человека и высших животных сохранили способность окислять глюкозу в анаэробных условиях, в результате чего при недостатке кислорода они имеют возможность получать энергию таким путем. Однако при этом образовавшаяся в анаэробных условиях молочная кислота, обладающая довольно большим запасом энергии, не выбрасывается в окружающую среду, а накапливается и мышцах. Из мышц она током крови доставляется в печень, где снова превращается в глюкозу. При поступлении в клетку достаточного количества кислорода часть молочной кислоты окисляется дальше до СО2 и Н2О. Превращение молочной кислоты. Образовавшаяся при анаэробном окислении глюкозы молочная кислота окисляется до СО2 и Н2О следующим образом. Сначала под действием фермента лактатдегидрогеназы, коферментом которой является НАД, она окисляется до пировиноградной кислоты:
которая затем под влиянием пируватдекарбоксилазы, представляющей собой сложный полиферментный комплекс, подвергается окислительному декарбоксилированию с образованием активной формы уксусной кислоты — ацетил- КоА:
где ТПФ — тиаминпирофосфат; ЛК — липоевая кислота; HSKoA — коэнзим А. В том случае, когда ткани хорошо снабжаются кислородом, пировиноградная кислота подвергается окислительному декарбоксилированию сразу, не восстанавливаясь до молочной кислоты. Восстановленный же кофермент НАД • Н + Н+, образовавшийся при окислении глицеральдегид-3-фосфата, передает водород через ферменты аэробного обмена (т.е. дыхательную цепь) на кислород, образуя воду. Превращение пировиноградной кислоты в ацетил-КоА является подготовительной, или переходной, стадией, благодаря которой углеводы через пировиноградную кислоту, а затем через ацетил-КоА включаются в новый этап — кислородное окисление. Другими словами, этот процесс — связующее звено между гликолизом и собственно дыханием. Однако уже в результате окислительного декарбоксилирования пировиноградной кислоты до ацетил-КоА высвобождается около 9 % всей энергии окисления глюкозы, т.е. больше, чем при гликолизе в целом, где высвобождается всего 5—7 % энергии. Если учесть 5—7 % энергии гликолиза и 9 % энергии окислительного декарбоксилирования пировиноградной кислоты, то всего выделяется 14—16 % энергии, аккумулированной в углеводах. Следовательно, остальные 84—86 % энергии сохраняется еще в молекуле уксусной кислоты. Цикл трикарбоновых кислот (цикл Кребса) представляет собой новый, более совершенный механизм окисления углеводов, выработанный у живых организмов с появлением на Земле кислорода. При помощи этого механизма происходит дальнейшее превращение уксусной кислоты в форме ацетил-КоА до СО2 и Н2О в аэробных условиях с высвобождением энергии. В связи с тем что первыми субстратами в процессе окисления уксусной кислоты являются трикарбоновые кислоты, а гипотезу о механизме этого окисления выдвинул X. А. Кребс, процесс назвали циклом трикарбоновых кислот, или циклом Кребса. Первой реакцией цикла является реакция конденсации ацетил-КоА со щавелевоуксусной кислотой, которую катализирует фермент цитратсинтаза. В результате образуется активная форма лимонной кислоты — цитрил-KoA:
Гидролизуясь, цитрил-КоА превращается в лимонную кислоту:
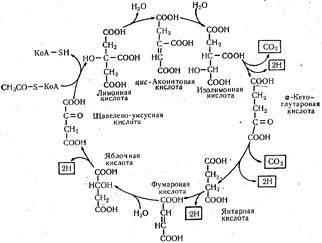
Последняя под действием фермента аконитатгидратаза превращается в цис-аконитовую кислоту, которая, присоединяя воду, превращается в изолимонную кислоту:
Изолимонная кислота далее окисляется путем отщепления двух атомов водорода, превращаясь в щавелевоянтарную. Этой реакцией начинается отщепление СО2 и первое окисление ацетил-КоА в трикарбоновом цикле. Щавелево-янтарная кислота, декарбоксилируясь, превращается в α -кетоглутаровую кислоту. Дегидрирование изолимон-ной и декарбоксилирование щавелево-янтарной кислот катализируется ферментом изоцитратдегидрогеназой с участием кофермента НАД+: Следующим этапом цикла трикарбоновых кислот является реакция окислительного декарбоксилирования α -кетоглутаровой кислоты, в результате которой образуется янтарная кислота. Этот процесс протекает в две стадии. Сначала α -кетоглутаровая кислота подвергается окислительному декарбоксилированию с образованием активной формы янтарной кислоты — сукцинил-КоА — и СО2. Эта реакция напоминает реакцию превращения пировиноградной кислоты до ацетил-КоА и катализируется также сложным полиферментным комплексом — α -кетоглутаратдегидрогеназой. В результате этой реакции происходит второе отщепление углекислого газа и дегидрирование уксусной кислоты, вступившей в цикл:
Образовавшаяся активная форма янтарной кислоты сукцинил-КоА, в отличие от ацетил-КоА, представляет собой макроэргическое тио-эфирное соединение, в котором аккумулирована энергия окисления α -кетоглутаровой кислоты. На следующей стадии эта энергия используется для образования ГТФ (гуанозинтрифосфорной кислоты) из ГДФ и неорганической фосфорной кислоты и запасается в фосфатных связях этого соединения. Реакция катализируется ферментом сукцинилтиокиназой:
Образовавшийся в результате этой реакции ГТФ взаимодействует с АДФ, в результате чего образуется АТФ: ГТФ + АДФ Синтез АТФ, сопряженный с окислением субстрата, является еще одним примером субстратного фосфорилирования. В дальнейшем ходе цикла трикарбоновых кислот происходит еще два дегидрирования. Янтарная кислота под действием сукцинатде-гидрогеназы с участием кофермента ФАД+ отщепляет два атома водорода и превращается в фумаровую кислоту, а ФАД+ восстанавливается до ФАД•Н2. Затем фумаровая кислота, присоединяя молекулу воды, образует яблочную кислоту (малат), которая при помощи малат-дегидрогеназы и кофермента НАД+ снова подвергается дегидрированию. При этом образуется щавелево-уксусная кислота, т.е. субстрат, с которого начался цикл трикарбоновых кислот:
Регенерированная щавелево-уксусная кислота может снова вступать в реакцию с новой молекулой ацетил-КоА, и процесс начнется в том же порядке. Общую схему цикла трикарбоновых кислот можно представить следующим образом: Цикл трикарбоновых кислот
(в рамках показаны конечные продукты окисления ацетил-КоА). Из приведенной схемы следует, что основная функция цикла Кребса заключается в дегидрировании уксусной кислоты. Если подвести баланс ферментативного дегидрирования одного цикла, можно легко подсчитать, что в результате реакций образуется восемь атомов водорода: шесть атомов используется для восстановления НАД+ и два — для восстановления ФАД+ сукцинатгидрогеназы. Суммарная реакция этого цикла описывается следующим уравнением: СН3СООН + 2Н2О → 2СО2 + 8Н, из которого следует, что четыре атома водорода принадлежат воде. Следовательно, остальные четыре образовались при дегидрировании уксусной кислоты, т.е. это весь водород, который был в составе ее молекулы. Одновременно с этим в виде оксида углерода (IV) дважды выделилось два атома углерода (один раз при декарбоксилировании щавелево-янтарной кислоты, второй — при декарбоксилировании α -кетоглутаровой кислоты), т.е. ровно столько, сколько их поступило в цикл в виде ацетальной группы. Из приведенного выше уравнения также следует, что в цикл не вовлекаются ни кислород, ни АТФ, ни неорганическая фосфорная кислота. Все эти метаболиты взаимодействуют в дыхательной цепи, куда вовлекаются неорганическая фосфорная кислота, отщепленные при дегидрировании атомы водорода и кислород, а в результате окислительного фосфорилирования образуется АТФ. Энергия для этого процесса выделяется в результате окислительно-восстановительных реакций при передаче атомов водорода и электронов от восстановленных форм НАД • Н2 и ФАД • Н2 на кислород. Процесс окислительного фосфорилирования подробно изложен в гл. 22. Напомним только, что на каждую пару электронов (пара атомов водорода) в дыхательной цепи путем окислительного фосфорилирования образуется три молекулы АТФ (одна при переносе атомов водорода от НАД • Н + Н+ к ФАД, вторая — при переносе пары электронов от цитохрома b к цитохрому с и третья — от цитохрома а3 к атому кислорода). Таким образом, каждая окислительная стадия превращения глюкозы до СО2 и Н2О, связанная с НАД, сопровождается образованием трех молекул АТФ, связанная с ФАД — образованием двух молекул АТФ. Энергетический баланс окисления углеводов. Сначала подведем итог энергетического баланса за счет дегидрирования уксусной кислоты в цикле Кребса. Как мы уже установили, в этом цикле происходит четыре дегидрирования, в результате которых образовались три восстановленные формы НАД, одна— ФАД и путем субстратного фосфорилирования синтезировалась одна молекула АТФ:
Таким образом, в цикле Кребса синтезируется в шесть раз больше АТФ, чем при гликолизе. Если учесть еще две восстановленные молекулы НАД, образовавшиеся при окислении молочной и пировиноград-ной кислот, то это составит еще 6 молекул АТФ, а в сумме— 18. Поскольку глюкоза распадается на две фосфотриозы, количество АТФ увеличивается в 2 раза и составит 36 молекул. Добавив к этому 2 молекулы АТФ, образовавшиеся в процессе гликолиза, получим общий баланс энергии, аккумулированной в мак-роэргических связях АТФ при окислении глюкозы до СО2 и Н2О: 36 + 2 = 38. Установлено, что полное окисление 1 моль глюкозы до СО2 и Н2О сопровождается выделением 2872 кДж. В 38 молекулах АТФ аккумулируется 1270—1560 кДж, т.е. приблизительно 50 % всей энергии, высвободившейся в процессе окисления. Следовательно, остальные 50 % энергии рассеиваются в организме в форме теплоты для поддержания соответствующей температуры. Из рассмотренных фаз окисления глюкозы исключительно важное значение имеет аэробная фаза. Если при анаэробном окислении, т.е. при образовании молочной кислоты, выделяется всего 197 кДж энергии, из которых 40 % аккумулируется в макроэргических связях двух молекул АТФ, то в аэробной фазе выделяется 2872 — 197 = = 2675 кДж, что составляет около 93 % всей энергии. Таким образом, основную массу энергии организм получает при дыхании. Апотомический путь окисления глюкозы. Наряду с циклом Кребса во многих клетках существует и другой путь расщепления глюкозы, называемый апотомическим, или пентозофосфатным. Экспериментально установлено, что в аэробных условиях в эритроцитах, печени, почках глюкоза может окисляться до 6-монофосфоглюконовой кислоты, причем фруктозо-1, 6-дифосфат в этом процессе не образуется. В результате такого окисления глюкозы образуется значительное количество пентоз. Этот путь был открыт советским биохимиком В. А. Энгельгардтом, а отдельные его этапы изучены О. Варбургом, Ф. Диккенсом, И. Д. Головацким и др. Пентозофосфатный путь не является главным путем окисления глюкозы. Основное его назначение состоит в том, чтобы снабжать клетки восстановленными формами НАДФ, необходимыми для биосинтеза жирных кислот, холестерина, пуриновых и пиримидиновых оснований, стероидов и др. Вторая функция этого пути заключается в том, что он поставляет пентозы, главным образом D-рибозу, для синтеза нуклеиновых кислот. Пентозофосфатный путь расщепления глюкозы суммарно можно выразить следующим уравнением: Глюкозо-6-монофосфат + 2 НАДФ+ → Рибозо-5-монофосфат + СО2 + 2 НАДФ·Н + Н+ + 2Н+. Пентозы, не использованные для биосинтеза нуклеиновых кислот и нуклеотидов, расходуются на биосинтез других соединений и регенерацию глюкозы. Биосинтез углеводов Существует два основных способа биосинтеза углеводов из относительно несложных метаболитов. Один из них заключается в восстановлении углекислого газа до глюкозы. Этот процесс, характерный для зеленых растений и называемый фотосинтезом, осуществляется за счет энергии солнечных лучей при помощи хлорофилла согласно следующему уравнению: СО2 + 2Н2О Улавливая солнечные лучи и преобразуя их энергию в энергию углеводов, зеленые растения обеспечивают сохранение и развитие жизни на Земле. В этом заключается, по словам К. А.Тимирязева, космическая роль зеленых растений как посредника между солнцем и всем живым на Земле. В последнее время работами группы ученых Института биохимии им. А. В. Палладина АН УССР под руководством академика М. Ф. Гулого показано, что ткани высших животных также способны фиксировать углекислый газ, хотя механизм фиксации его отличается от такового у фотосинтезирующих клеток. Он заключается в наращивании углеродного скелета оксидом углерода (IV) таких субстратов, как кетокислоты, жирные кислоты, аминокислоты и др. В печени, почках и скелетных мышцах человека и высших животных существует другой путь биосинтеза углеводов, называемый глюконеогенезом. Это синтез глюкозы из пировиноградной или молочной кислоты, а также из так называемых гликогенных аминокислот, жиров и других предшественников, которые в процессе метаболизма могут превращаться в пировиноградную кислоту или метаболиты цикла трикарбоновых кислот. Глюконеогенез— это путь, обратный гликолизу. Однако в этом пути есть три стадии, которые в энергетическом отношении не могут быть использованы при превращении пировиноградной кислоты в глюкозу. Эти три стадии гликолиза заменены «обходными» реакциями с меньшей затратой энергии. Первой обходной реакцией является превращение пировиноградной кислоты в фосфоенолпировиноградную. Поскольку расщепление глюкозы происходит в митохондриях, а синтез — в цитоплазме, на первом этапе митохондриальная пировиноградная кислота превращается сначала в щавелево-уксусную. Катализирует это превращение фермент пируваткарбоксилаза, активируемая ацетил-КоА с участием АТФ. Образовавшаяся щавелево-уксусная кислота восстанавливается затем с участием НАД • Н +Н+ в яблочную: Пировиноградная кислота + СО2 Яблочная кислота диффундирует в цитоплазму, окисляется цитоплазматической малатдегидрогеназой с образованием цитоплазматической щавелево-уксусной кислоты, из которой образуется фосфоенолпировиноградная кислота. Эту реакцию катализирует фосфоенолпируваткарбоксикиназа. Донором фосфорной кислоты служит ГТФ: Яблочная кислота Далее следует целая серия обратных реакций, заканчивающихся образованием фруктозо-1, 6-дифосфата. Превращение фруктозо-1, 6-дифосфата в фруктозо-6-фосфат— вторая необратимая реакция гликолиза. Поэтому она катализируется не фосфофруктокиназой, а фруктозодифосфатазой. Этот фермент катализирует необратимый гидролиз 1-фосфатной группы: Фруктозо-1, 6-дифосфат + Н2О → Фруктозо-6-фосфат + Н3РО4. На следующей (обратимой) стадии биосинтеза глюкозы фруктозо-6-фосфат превращается в глюкозо-6-фосфат под действием фосфоглюко-изомеразы гликолиза. Расщепление глюкозо-6-фосфата до глюкозы — третья необратимая реакция, которая не осуществляется путем обращения гексокиназой. Свободная глюкоза образуется при помощи глюкозо-6-фосфатазы, катализирующей реакцию гидролиза: Глюкозо-6-фосфат +Н2О → Глюкоза + H3PO4. В большинстве клеток глюкозо-6-фосфат, образующийся в процессе гликогенолиза, используется как предшественник для биосинтеза олиго- и полисахаридов. Большую роль в биосинтезе этих сложных сахаров играет соединение уридинфосфоглюкоза, которая выполняет роль промежуточного переносчика глюкозы. При биосинтезе гликогена, например, глюкозо-6-фосфат, превратившись в глюкозо-1-фосфат под действием фосфоглюкомутазы, взаимодействует с уридинтрифосфорной кислотой (УТФ) — соединением, аналогичным АТФ, в которое вместо аденина входит азотистое основание урацил. В результате этого взаимодействия при помощи глюкозо-I -фосфатуридилтрансферазы образуется уридилдифосфоглюкоза: Глюкозо-1-фосфат + УТФ На заключительном этапе биосинтеза гликогена в реакции, катали-зируемой гликогенсинтетазой, остаток глюкозы с УДФ-глюкозы переносится на концевой остаток глюкозы амилазной цепи с образованием 1, 4-гликозидной связи (см. гл. 16). Ветвление гликогена путем образования 1, 6-связей завершается амило-1, 4-1, 6-трансглюкозидазой. Биосинтез гликогена осуществляется не только из глюкозо-6-фосфата, образовавшегося путем глюконеогенеза. Как уже отмечалось выше, для его биосинтеза используется также часть глюкозы после всасывания. Синтез гликогена, как процесс образования подвижного резерва углеводов в организме, имеет большое биологическое значение. Ведущая роль в этом принадлежит печени. Благодаря синтезу и отложению гликогена в печени поддерживается постоянная концентрация глюкозы в крови и других тканях, а также предотвращаются потери ее с мочой при употреблении пищи, особенно углеводной. Кроме того, отложение гликогена в печени способствует постепенному использованию углеводов в зависимости от условий существования организма. Использованию глюкозы для синтеза гликогена предшествует образование глюкозофосфорных эфиров. Сначала образуется глюко-зо-6-монофосфат. Источником энергии и донатором фосфата является АТФ. Катализирует эту реакцию гексокиназа. Под действием фермента фосфоглюкомутазы глюкозо-6-монофосфат превращается в глюкозо-1-монофосфат:
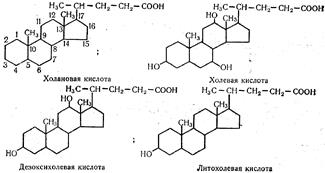
Дальнейшее превращение глюкозо-1-монофосфата до гликогена протекают уже знакомым нам путем. Глава 24. ОБМЕН ЛИПИДОВ Липиды представляют собой большую группу органических соединений. Все они различаются по своему химическому составу и структуре, но обладают одним общим для них свойством — нерастворимостью в воде. В связи с тем что ферменты, действующие на эти органические соединения, водорастворимы, расщепление и всасывание липидов в пищевом канале характеризуются некоторыми особенностями. Наличие же липидов различной структуры обусловливает различные пути их расщепления и синтеза. Остановимся на обмене жиров, фосфатидов и стеридов, имеющих наиболее важное биологическое значение. Обмен липидов, как и углеводов, — многоступенчатый процесс, который состоит из пищеварения, всасывания, транспортирования липидов кровью, внутриклеточного окисления и биосинтеза. Переваривание липидов Переваривание триглицеридов. Триглицериды, или нейтральные жиры, являются концентрированными источниками энергии в организме. При окислении 1 г жира высвобождается около 38, 9 кДж энергии. Являясь гидрофобными соединениями, жиры резервируются в компактной форме, занимая сравнительно мало места в организме. Вместе с пищей в организм человека ежесуточно поступает до 70 г жиров растительного и животного происхождения. По своей химической природе они являются главным образом триглицеридами. Расщепление жиров происходит при помощи ферментов, называемых липазами. Слюна не содержит таких ферментов, поэтому в ротовой полости жиры никаким изменениям не подвергаются. В желудке активность липазы очень слабая. Это связано с тем, что в желудке реакция среды сильнокислая (рН = 1, 5-2, 5), в то время как оптимум действия липазы находится при рН = 7, 8 = 8, 1. В связи с этим в желудке переваривается всего 3-5 % поступающих жиров. Переваривание жиров в желудке происходит только у новорожденных и детей грудного возраста. Это связано с тем, что рН среды в желудке новорожденных составляет 5, 6, а в этих условиях липаза проявляет большую активность. Кроме того, жир материнского молока, которое является основным продуктом питания детей в этот период, находится в сильно эмульгированном состоянии, а само молоко содержит липолитический фактор, принимающий участие в переваривании жиров. Однако желудок все же играет определенную роль в процессе переваривания жиров у взрослых. Он регулирует поступление жира в кишки и переваривает белки, освобождая таким путем жир из липопротеидных комплексов пищи. Основным местом переваривания жиров является двенадцатиперстная кишка и отделы тонкой кишки. Поскольку жиры нерастворимы в воде, а ферменты, расщепляющие их, являются водорастворимыми соединениями, необходимым условием для гидролитического расщепления жиров на составные части является их диспергирование (дробление) с образованием тонкой эмульсии. Диспергирование и эмульгирование жира происходит в результате действия нескольких факторов: желчных кислот, свободных высших жирных кислот, моно- и диглицеридов, а также белков. Этому способствуют также перистальтика кишок и постоянно образующийся углекислый газ, который выделяется при взаимодействии кислых компонентов пищи, поступающих из желудка, с карбонатами кишок, создающими щелочную среду. Образовавшийся углекислый газ «пробулькивает» через пищевые массы, участвуя таким образом в диспергировании жира. Нейтрализации содержимого желудка способствует также поступление в просвет тонкой кишки желчи, обладающей щелочным характером.) Желчь — вязкая жидкость светло-желтого цвета со специфическим запахом, горькая на вкус. В состав желчи входят желчные кислоты. желчные пигменты, продукты распада гемоглобина, холестерин, лецитин, жиры, некоторые ферменты, гормоны и др. Желчь способствует перистальтике тонкой кишки, оказывает бактериостатическоедействие на ее микрофлору. С желчью выделяются из организма яды. Она является также активатором липолитических ферментов и повышает проницаемость стенки кишок. Главной составной частью желчи являются желчные кислоты. Они образуются в печени из холестерина и находятся в желчи как в свободном, так и в связанном состоянии, а также в виде натриевых солей. В желчи человека содержится в основном три желчных кислоты Основную массу составляют холевая (3, 7, 12-тригидроксихола-новая) и дезоксихолевая (3, 12-дигидроксихолановая), небольшую часть — литохолевая (3-гидроксихолановая) кислоты, которые являются производными холановой кислоты:
Холевая кислота может находиться в желчи также в связанном состоянии в виде парных соединений с глицином и производным цистеина таурином — соответственно гликохолевой и таурохолевой кислот:
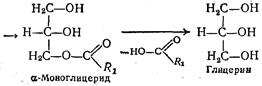
Благодаря наличию желчных кислот происходит снижение поверхностного натяжения липидных капель, что способствует образованию очень тонкой и устойчивой эмульсии диаметр частиц которой составляет около 0, 5 мкм. Образованию эмульсии способствуют также моноглицериды и высшие жирные кислоты. Эмульгирование жира приводит к колоссальному увеличению поверхности соприкосновения липазы с водным раствором. Таким образом, чем тоньше эмульсия жиров, тем лучше и быстрее они расщепляются липазой. Кроме того, в виде тонкой эмульсии жиры могут даже всасываться стенкой кишок непосредственно, не расщепляясь на составные части. В присутствии желчных кислот под действием липазы в просвете тонкой кишки происходит гидролитическое расщепление жиров. В результате этого образуются продукты частичного и полного расщепления жиров — моно- и диглицериды, свободные высшие жирные кислоты и глицерин:
Здесь же содержится и часть нерасщепленного жира в виде очень тонкой эмульсии. Все эти продукты в дальнейшем всасываются стенкой кишок. В этой смеси триглицериды составляют около 10 % , моно- идисахариды — также 10 % , а основная масса — около 80 % — продукты полного расщепления жиров— глицерин и высшие жирные кислоты. Переваривание фосфоглицеридов. Основным местом переваривания фосфатидов также является двенадцатиперстная кишка. Эмульгирование этих липидов происходит под влиянием тех же веществ, что и три-глицеридов. Однако гидролитическое расщепление фосфатидов осуществляется под действием фосфолипаз А, В, С и D. Каждый фермент действует на определенную сложноэфирную связь фосфолипида. Гидролитическое расщепление, например, лецитина происходит следующим образом:
Такому полному расщеплению подвергается незначительная часть фосфатидов, поскольку его промежуточные продукты хорошо растворимы в воде и легко всасываются стенкой кишок. К тому же фосфогли-цериды легко образуют эмульсии, которые также могут всасываться кишечной стенкой. Переваривание стеридов. Стериды, входящие в состав пищи, эмульгируются под влиянием тех же факторов, что и жиры, после чего подвергаются гидролитическому расщеплению до свободных стеринов и высших жирных кислот. Этот процесс осуществляется под действием фермента холестеринэстеразы. Всасывание липидов Врезультате пищеварения жиров, фосфатидов, стеридов в просвете тонкой кишки образуется значительное количество продуктов их частичного и полного гидролитического расщепления: моно- и диглицериды, высшие жирные кислоты, стерины, азотистые основания, фосфорная кислота. Содержится также небольшое количество триглицеридов, находящихся в тонкоэмульгированном состоянии. Все эти продукты всасываются стенкой тонкой кишки. Такие продукты расщепления, как жирные кислоты и холестерин, плохо растворяясь в воде, образуют с желчными кислотами водорастворимые комплексы— так называемые холеиновые кислоты. Эти кислоты легко проникают в эпителиальные клетки стенки кишок, где расщепляются на составные части. Освобожденные желчные кислоты возвращаются в просвет кишок и снова используются для транспортирования нерастворимых в воде продуктов расщепления жиров. Часть продуктов расщепления (глицерин, глицеринфосфорная кислота, азотистые основания) хорошо растворимы в воде и легко проникают в эпителиальные клетки. Фосфорная кислота всасывается в клетки эпителия стенки тонкой кишки в виде натриевых и калиевых солей. В основе всасывания липидов лежит ряд сложных физико-химических и биологических процессов, для осуществления которых затрачивается энергия макроэргических связей АТФ. В эпителиальных клетках слизистой оболочки кишок из всосавшихся продуктов гидролитического расщепления снова синтезируются липиды. Однако этот ресинтез приводит к образованию специфических жиров, характерных для данного организма. Для образования нейтральных жиров используются высшие жирные кислоты, глицерин, моно- и диглицериды. Одновременно происходит и синтез фосфатидов, для которых используются главным образом глицеринфосфорная кислота, глицериды и диглицериды, а также в небольшом количестве моноглицериды. Из холестерина и высших жирных кислот образуются стериды. В эпителиальных клетках стенки кишок из синтезированных липидов, а также капель всосавшихся триглицеридов, витаминов (A, D, Е, К) ибелков образуются комплексы размером 150—200 нм, называемые хило микронами. Внутреннее содержимое хиломикрона, представленное образовавшимися различного рода липидами, главным образом триглицеридами, окружено наружной белковой оболочкой, благодаря которой хиломикроны хорошо растворяются в воде. Хило-микроны диффундируют сначала в межклеточную жидкость, затем в лимфатические капилляры и в конце концов попадают в кровяное русло, где под действием гепарина распадаются на мелкие частицы. С током крови они разносятся по всему организму и откладываются в резерв в жировых депо — подкожной и околопочечной клетчатке, сальнике, брыжейке, мышечной ткани. Часть жиров крови используется для пластических целей, как источник химической энергии и т.д. Таким образом, хиломикроны являются переносчиками образовавшихся в эпителиальных клетках тонкой кишки липидов. При этом они транспортируют в крови главным образом триглицериды. Наряду с хиломикронами существуют и другие формы транспорта липидов кровью, например α - и β -липопротеиды. Их молекулы представляют собой сложные комплексы липидов с белками. α -Липопро-теиды являются основными транспортными формами фосфатидов, β -липопротеиды— переносчиками холестерина и его эфиров. Наиболее подвижной формой липидов являются свободные высшие жирные кислоты. Важная роль в активном транспортировании липидов принадлежит форменным элементам крови. Эритроциты, например, участвуют в переносе фосфатидов и холестерина, лейкоциты— триглицеридов. Большая роль в обмене липидов принадлежит жировым депо. Исследования показали, что в жировых депо откладывается не только вновь синтезированный в организме специфически видовой жир, но и в небольших количествах чужеродный, т.е. входящий в состав пищи. Опыты, проведенные на голодающих собаках, показали, что пищевые жиры после всасывания поступают сначала в жировые депо, из которых переходят в плазму крови. Таким образом, жировая ткань не является пассивным депо жиров, состав ее постоянно обновляется за счет липидов, всасывающихся из кишок или синтезируемых в организме. |
Последнее изменение этой страницы: 2017-03-14; Просмотров: 4997; Нарушение авторского права страницы








 ГДФ + АТФ.
ГДФ + АТФ.
 1/6С6Н12О6 + О2 + Н2О.
1/6С6Н12О6 + О2 + Н2О. Щавелево-уксусная кислота
Щавелево-уксусная кислота  Яблочная кислота.
Яблочная кислота.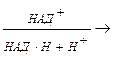 Щавелево-уксусная кислота
Щавелево-уксусная кислота  Фосфоенолпировиноградная кислота.
Фосфоенолпировиноградная кислота. УДФ-глюкоза+Фн.
УДФ-глюкоза+Фн.
 Натриевая соль гликохолевой кислоты
Натриевая соль гликохолевой кислоты Натриевая соль таурохолевой кислоты
Натриевая соль таурохолевой кислоты