|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Роль высокоэнергетических соединений в клеточном метаболизме
5.1 Специализированное применение двух универсальных форм энергии АТФ участвует в реакциях, протекающих в цитоплазме, то есть в форме АТФ энергией обеспечиваются все процессы, протекающие в водной среде. К числу последних относится большинство биосинтетических реакций. Помимо этого АТФ служит источником энергии для протекания ряда мембранзависимых процессов. Энергия протонного градиента связана исключительно с мембранами, которые являются необходимым компонентом для его образования. Поэтому энергией в форме протонного градиента (∆ μ н+) могут обеспечиваться только процессы, локализованные на мембране. Таким образом, у протонного градиента (∆ μ н+) более узкая область приложения. В то же время использование клеткой энергии в форме протонного градиента (∆ μ н+) имеет определенные преимущества: протонный градиент (∆ μ н+) в форме его электрической составляющей — более удобная форма энергии для внутри- и межклеточной транспортировки. Скорость переноса энергии посредством диффузии АТФ в цитоплазме значительно медленнее, чем скорость передачи протонного градиента (∆ μ н+) по мембранам. Энергия в форме протонного градиента (∆ μ н+) не содержится в виде определенных порций, как это имеет место в молекуле АТФ. При гидролизе макроэргической фосфатной связи АТФ освобождается определенное количество энергии (∆ G0 = -31, 8 кДж/моль). Если для сопряженного эндергонического процесса требуется меньшее количество энергии, остальная часть рассеивается в виде тепла. При использовании энергии в форме трансмембранного потенциала потерь, обусловленных запасанием энергии в виде порций, не происходит. С этим связано и еще одно преимущество при использовании клеткой энергии в форме протонного градиента (∆ μ н+), а именно-не существует нижнего порога для его образования. Роль других соединений с богатыми энергией связями
Слайд №33 Эти соединения перечислены, в таблице 3 вместе с примерами реакции активации, в которых они участвуют. Роль пиридиновых нуклеотидов в метаболизме Как и все остальные реакции окисления, биологическое окисление органических метаболитов представляет собой удаление электронов. В большинстве случаев на каждой стадии окисления метаболита происходит удаление двух электронов и одновременно потеря двух протонов. Этот процесс эквивалентен отнятию двух атомов водорода и называется дегидрированием. Наоборот, восстановление какого-либо метаболита представляет собой присоединение двух электронов и двух протонов и, следовательно, его можно рассматривать как гидрирование. Посредниками в реакциях биологического окисления и восстановления чаще всего являются два пиридиновых нуклеотида: никотинамидадениндинуклеотид (НАД) и никотинамидадениндинуклеотидфосфат (НАД Ф). Оба эти пиридиновых нуклеотида легко могут претерпевать обратимое окисление и восстановление никотинамидной группы. Окисленная форма пиридиновых нуклеотидов содержит на один атом водорода меньше, чем восстановленная форма; кроме того, она имеет положительный заряд, позволяющий ей акцептировать при восстановлении второй электрон. Обратимые реакции окисления — восстановления НАД и НАДФ можно записать следующим образом: Слайд№34
НАД+ + 2Н ↔ НАД Н + Н+ (окислен форма) (восстановл. форма) НАДФ+ + 2Н ↔ НАДФ Н + Н+.
Функция пиридиновых нуклеотидов в метаболизме состоит в переносе восстановительного потенциала или в его образовании. Каждая форма клеточной энергии может быть использована для осуществления химической, механической и осмотической работы, между ними существует определенное " разделение труда". Например, большинство биосинтезов обеспечивается энергией АТФ, активный транспорт — энергией протонного градиента (∆ μ н+). Из этого следует, что клетке необходимо всегда иметь определенное количество энергии в той и другой легко мобилизуемой форме. Это может быть одной из причин существования в клетке двух взаимосвязанных энергетических пулов (резервуаров), между которыми при необходимости легко может осуществляться перекачка энергии (см. рисунок 3). Емкость обоих энергетических пулов невелика. Например, величина протонного градиента (∆ μ н+) поддерживается на уровне 200-250мВ. Внутриклеточная концентрация АТФ составляет около 2мМ. Это также указывает на каталитическую роль АТФ в клетке. Подсчитано, что для удвоения клеточной массы молекула АТФ должна около 10000 раз участвовать в реакциях гидролиза и синтеза.
5.3 Энергетические затраты микробной клетки В растущей бактериальной культуре потребление энергии в первую очередь связано с процессами биосинтеза веществ, из которых состоит клетка. Количество энергии, необходимое для биосинтетических целей, в большой степени зависит от состава среды культивирования. Теоретически рассчитано, что при выращивании культуры бактерий гетеротрофов в среде с минеральными солями и глюкозой в качестве единственного источника углерода 1 моль АТФ затрачивается для синтеза 27г вещества клеток. Если же единственным источником углерода служит двуокись углерода (хемотрофы), тогда использование того же количества АТФ приведет к синтезу только 5г вещества клеток. Кроме энергетических затрат на биосинтетические процессы, связанных с ростом, определенная часть клеточной энергии всегда тратится на процессы, не связанные непосредственно с ростом. Последние получили название процессов поддержания жизнедеятельности. К специфическим функциям поддержания жизнедеятельности относятся: обновление клеточного материала, осмотическая работа, обеспечивающая поддержание концентрационных градиентов между клеткой и внешней средой, подвижность клетки и др. Энергию, расходующуюся на осуществление перечисленных функций, обозначают как энергию поддержания жизнедеятельности.
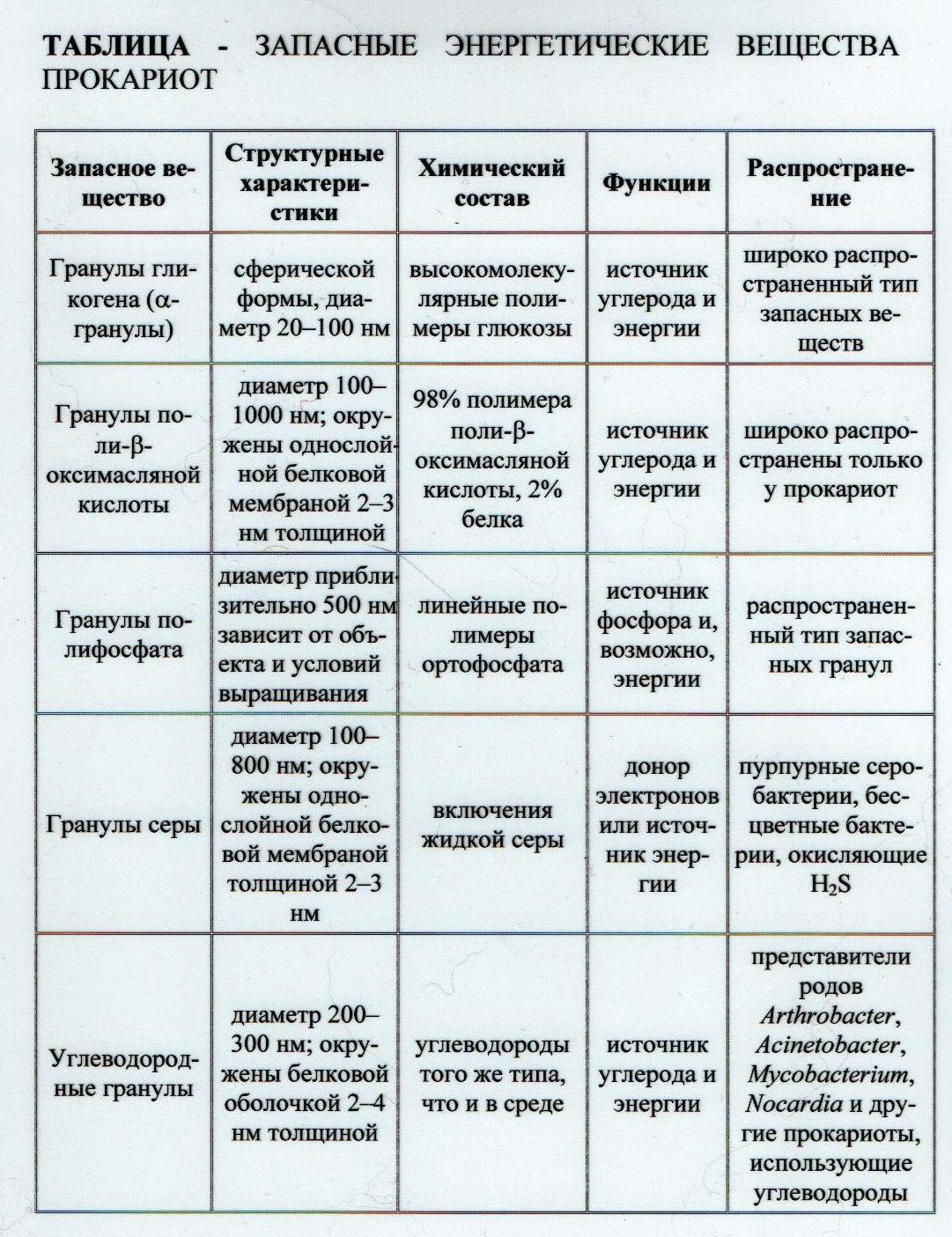
5.4 Консервирование энергии в микробной клетке Возможны такие условия, когда клетка запасает энергии больше, чем тратит. В этом случае она сталкивается с проблемой консервирования энергии. В молекулах АТФ энергия не хранится в течение длительного времени. Средняя продолжительность " жизни" молекул АТФ составляет около 1/3с. Энергия в форме протонного градиента (∆ μ н+) также не может накапливаться. Движение Н+ против градиента возможно только до достижения определенного уровня, после которого возникшая разность концентраций и электрических зарядов будет тормозить поступление ионов водорода против градиента. Таким образом, в молекулах АТФ и в виде протонного градиента (∆ μ н+) энергия находится в мобильной форме, призванной обеспечивать все идущие в настоящий момент энергозависимые процессы. Проблема консервирования энергии решена прокариотами путем синтеза восстановленных высокополимерных молекул. Эти запасные энергетические вещества представлены полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы (таблица 4). Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобкое вещество — гранулеза. Последняя -специфический запасной полисахарид анаэробных споровых бактерий группы клостридий. Названные полисахариды построены из остатков глюкозы. В неблагоприятных условиях они используются в качестве источника углерода и энергии.
Липиды накапливаются в виде гранул, резко преломляющих свет и поэтому хорошо различимых в световой микроскоп. Запасным веществом такого рода является полимер β -оксимасляной кислоты, накапливающийся в клетках многих прокариот. Другой широко распространенный тип запасных веществ многих прокариот — полифосфаты, содержащиеся в гранулах, называемых волютиновыми, или метахроматиновыми, зернами. Используются клетками как источник фосфора. Полифосфаты содержат макроэргические связи и, таким образом, являются депо энергии, хотя считается, что их роль как источника энергии незначительна. Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в клетках молекулярной серы. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды оказывается исчерпанным. Для аэробных тионовых бактерий, окисляющих сероводород (H2S), сера служит источником энергии, а для анаэробных фотосинтезирующих серобактерий она является донором электронов. Молекулы запасных энергетических веществ представлены в виде высокомолекулярных полимерных молекул, плотно упакованы в гранулах и часто окружены белковой оболочкой. В таком виде они находятся в осмотически неактивном состоянии, что очень важно для клетки, так как в противном случае сосредоточение в цитоплазме большого числа молекул осмотически активных веществ оказало бы на клетку отрицательное действие. Лекция №1.3
Тема: «Дыхание, брожение и фотосинтез микроорганизмов. Методы исследования обмена веществ у микроорганизмов»
Вопросы: |
Последнее изменение этой страницы: 2017-04-12; Просмотров: 630; Нарушение авторского права страницы