|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Компонентный состав бактериальной клетки
Слайд №13 Клетки бактерий содержат от 70 до 90% воды. Основную массу сухих веществ, на долю которых приходятся остальные 10-30%, составляют белки, нуклеиновые кислоты, липиды и полисахариды. Несколько процентов сухого вещества клеток приходится на низкомолекулярные органические вещества и соли (таблица 1) Макромолекулы, составляющие основную массу сухих веществ клетки это полимеры, построенные из мономерных частиц. Образованию полимеров предшествует синтез составляющих их мономеров. В случае полисахаридов — это различные моносахара, нуклеиновых кислот — рибо- и дезоксирибонуклеотиды, белков — аминокислоты. Исключение составляют липиды, не являющиеся полимерами, так как молекулы в них не соединены между собой ковалентными связями. Слайд № 14 Липиды — гетерогенный класс клеточных компонентов, выделенный на основании их растворимости в неполярных растворителях (эфир, бензол, хлороформ) и нерастворимости в воде. К ним относят жиры, фосфолипиды, стероиды, изопреноиды и поли-β -оксибутират. Липиды подразделяют на два больших класса: липиды, содержащие жирные кислоты, связанные эфирной связью и липиды, состоящие из повторяющихся пятиуглеродных изопреновых фрагментов. Большое значение имеют сложные липиды. Они представляют собой сложный эфир глицерола, в котором две спиртовые группы этерифицированы жирными кислотами, а третья - остатком фосфорной кислоты или сахаром. Углеводные полимеры построены на основе повторяющихся единиц одного, двух или более типов углеродных соединений, например, полисахарид гликоген, построенный из остатков глюкозы, или пептидогликан клеточной стенки, образованный чередованием N-ацетилглюкозамина и N-ацетилмурамовой кислоты. Полимерные молекулы белков и нуклеиновых кислот синтезируются на матрице, которая и определяет последовательность составляющих их мономеров. Возможности для синтеза разнообразных по функциям и структуре клеточных метаболитов реализуются на стадии сборки полимеров путем различных сочетаний аминокислот. В основе огромного количества разных белков лежат комбинации из 20 аминокислот, а чтобы зашифровать весь объем генетической информации одной клетки или многоклеточного организма оказалось достаточным комбинации из 4 нуклеотидов.
2.2 Биосинтез аминокислот и белков 2.2.1 Аминокислоты. Большинство микроорганизмов способны самостоятельно синтезировать de novo все двадцать аминокислот, из которых строятся белки. Основное назначение источников азота, поступающих в бактериальную клетку, это формирование аминных (NH2) и иминных (NH) групп в молекулах аминокислот, нуклеотидов, гетероциклических оснований и других химических компонентов. При этом азотсодержащие вещества являются сырьём для пластического обмена но могут включаться и в энергетический метаболизм. Наиболее доступные минеральные источники азота в природе — аммонийный ион (NH4+) и аммиак (NH3), легко проникают в клетки и просто трансформируются в амино- и иминогруппы. Слайд №15 Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Аминогруппы вводятся путем прямого аминирования или трансаминирования. Перевод неорганического азота в органические соединения происходит всегда через аммиак. Нитраты, нитриты и молекулярный азот предварительно восстанавливаются до аммиака (ассимиляционная нитратредукция) и только после этого включаются в состав органических соединений (рисунок 3, а, б, в). Рисунок 3 Важнейшие пути ассимиляции азота в микробной клетке.\
Ионы аммония, содержащиеся в питательной среде, непосредственно поглощаются клетками (а). Ионы нитрата при ассимиляционной нитратредукции (б), а молекулярный азот (N2) при фиксации азота (в) восстанавливаются до ионов аммония. В органические соединения аммонийный азот переводится либо при участии АТФ путем образования глутамина, либо без затраты АТФ путем прямого восстановительного аминирования 2-оксоглутарата или пирувата. Лишь немногие из аминокислот образуются в результате прямого аминирования свободными ионами NH4+. Большинство аминокислот получают свою аминогруппу от одной из первичных аминокислот в результате трансаминирования. Из свободных аминокислот в цитоплазме количественно преобладает глутаминовая кислота (больше половины всего «пула» аминокислот). Слайд №16.Таким образом, глутаминовая кислота и глутамин прямо или косвенно служат донорами амино- и амидогрупп при синтезе практически всех аминокислот и других азотсодержащих органических соединений. Аспарагин используется только для синтеза белковых молекул. Во все остальные аминокислоты азот вводится посредством реакций переаминирования, катализируемых соответствующими аминотрансферазами, при этом во всех реакциях одним из компонентов является глутаминовая кислота: глутаминовая кислота + щавелевоуксусная кислота —> аспарагиновая кислота + α -кетоглутаровая кислота. Слайд №17.Еще одним путем включения азота аммиака в состав органических соединений является реакция, приводящая к образованию карбамоилфосфата:
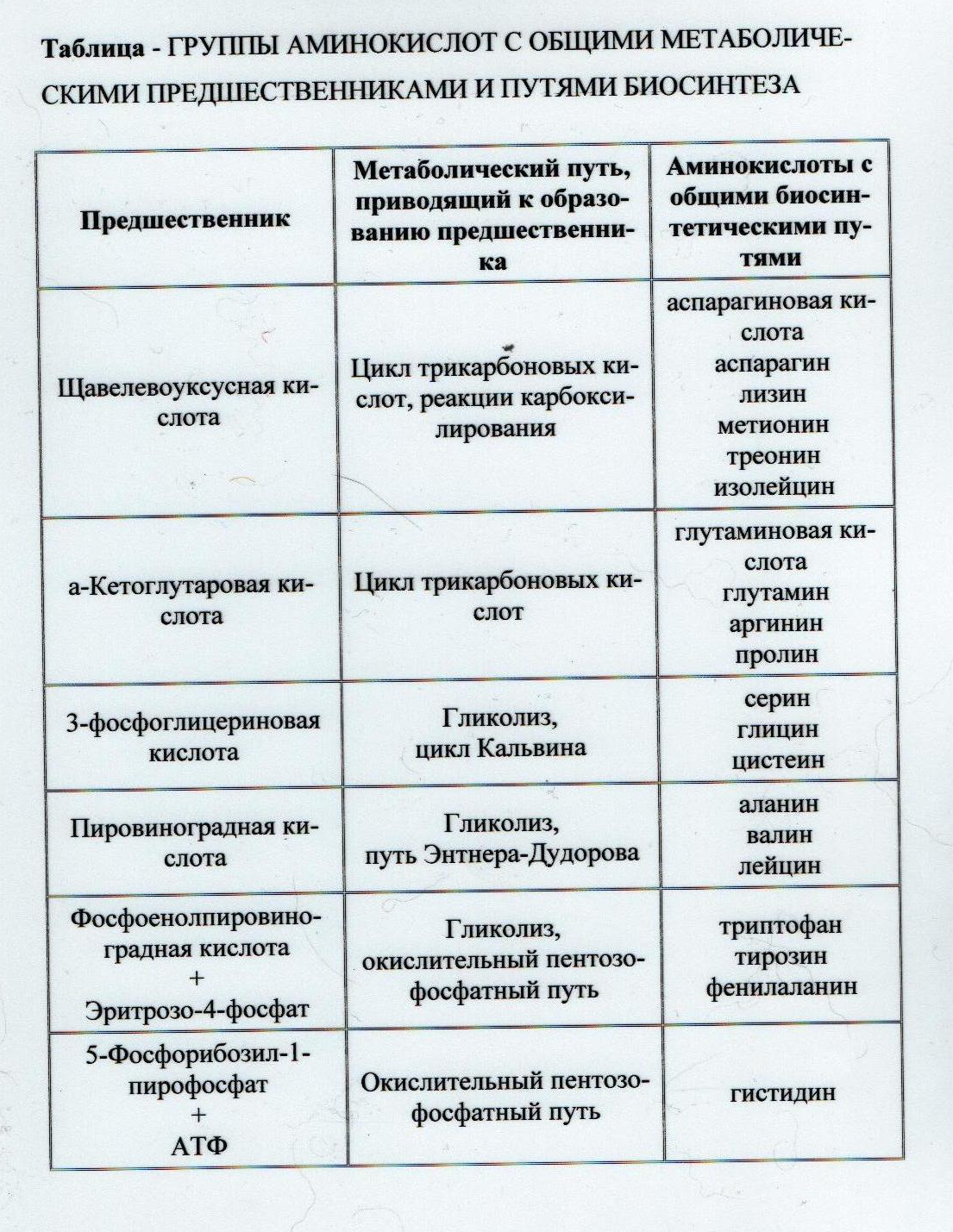
Дальнейшее использование азота карбамоилфосфата происходит по двум путям: для синтеза пиримидинов и аргинина. Слайд №18 У многих микроорганизмов хорошо изучены пути синтеза всех двадцати аминокислот. Особенностью биосинтеза аминокислот является использование общих биосинтетических путей. Так, 19 из 20 аминокислот, входящих в состав белков, можно по способу их биосинтеза разделить на 5 групп. Только одна аминокислота (гистидин) образуется по отдельному биосинтетическому пути. В обобщённом виде группирование синтезируемых аминокислот по признаку единства их метаболических предшественников и путей биосинтеза представлено в таблице 2. 2.2.2 Пептиды и белки. Бактериальная клетка способна синтезировать несколько тысяч различных белков, каждый из которых содержит в среднем 200 аминокислотных остатков. Информация, направляющая синтез этих белков, закодирована в ДНК. Синтез полипептидной цепи происходит в цитоплазме клетки на рибосомах в сочетании с молекулой матричной (мРНК) или информационной РНК (иРНК), которая синтезируется на матрице ДНК в процессе транскрипции. Информация, содержащаяся в молекулах мРНК, транслируется в полипептидную цепь при участии особого класса молекул РНК, известных как тРНК. Многофункциональность тРНК позволяет им присоединяться к определённым аминокислотам, связываться с рибосомой и узнавать определённые последовательности из трёх нуклеотидов (кодон) в составе мРНК. Узнаваемый кодон соответствует конкретной аминокислоте; нужная аминокислота «подаётся» при помощи узнающей её молекулы тРНК к концу растущей полипептидной цепочки. Так образуется будущая молекула белка.
2.3 Биосинтез нуклеотидов и нуклеиновых кислот 2.3.1 Пуриновые и пиримидиновые нуклеотиды. Из мононуклеотидов построены нуклеиновые кислоты (РНК, ДНК) клеток. Кроме того, мононуклеотиды входят в состав многих коферментов и участвуют в осуществлении различных каталитических функций. Центральное место в биосинтезе мононуклеотидов занимает синтез пуриновых и пиримидиновых азотистых оснований. Многие прокариоты способны использовать содержащиеся в питательной среде готовые пуриновые и пиримидиновые основания, их нуклеозиды и нуклеотиды. Слайд №19 Образование пуриновых и пиримидиновых нуклеотидов de novo из низкомолекулярных соединений происходит независимыми друг от друга путями. Источник рибозилфосфатной части всех нуклеотидов — 5-фосфорибозил-1-пирофосфат — образуется после взаимодействия рибозо-5-фосфата с АТФ. Дезоксирибонуклеотиды образуются путём восстановления соответствующих рибонуклеотидов. Пуриновые нуклеотиды построены на основе фосфорибозилпирофосфата, с которого и начинается их синтез. Девятичленное пуриновое кольцо синтезируется последовательным присоединением аминогрупп и мелких углеродсодержаших групп, причём все интермедиаты биосинтеза — рибонуклеотиды. Пиримидиновые нуклеотиды формируются при последовательных превращениях карбоксилсодержащих интермедиатов, начиная с карбамоилфосфата. Рибозофосфатный остаток присоединяется только после того, как шестичленное пиримидиновое кольцо полностью синтезировано из аспарагиновой кислоты и карбамоилфосфата. 2.3.2 Нуклеиновые кислоты. Большинство синтезируемых нуклеотидов используются для синтеза РНК и ДНК, небольшая часть используется для синтеза коферментов и богатых энергией соединений. Основная часть бактериальной ДНК находится в виде двухцепочечной кольцевой молекулы — бактериальной хромосомы. В процессе репликации хромосома с высокой точностью удваивается, что обеспечивает передачу генетической информации дочерним клеткам. Синтез ДНК из субстратов-мономеров четырёх дезоксинуклеозидтрифосФатов (дАТФ, дГТФ, дЦТФ и дТТФ), катализируют ДНК-полимеразы. При этом на каждой одиночной цепи ДНК происходит синтез новой комплементарной цепи: ДНК-полимеразы встраивают нуклеотиды в соответствии с правилами водородного связывания (Гуанин против Цитозина, Аденин против Тимина). Процесс удвоения клеточной ДНК называют репликацией. Если цепь ДНК служит матрицей, на которой полимеризуется цепь РНК (мРНК), то этот процесс называют транскрипцией.
2.4 Биосинтез олиго- и полисахаридов На долю внутриклеточных олиго- и полисахаридов может приходиться до 60% сухой массы бактериальной клетки, тогда как количество синтезируемых внеклеточных полисахаридов может во много раз превышать массу бактерии. При отсутствии сахаров в окружающей среде бактерии синтезируют их из доступных источников углерода. Например, бактерии, растущие на средах, содержащих трёхуглеродные соединения, синтезируют гексозофосфаты из пирувата с помощью реакций, характерных для фруктозодифосфатного пути. Бактериальные олиго- и полисахариды образуются путём присоединения к акцепторным молекулам остатков сахаров из нуклеозиддифосфосахаров. Например, трегалоза, содержащая два остатка глюкозы, образуется из уридиндифосфатглюкозы и глюкозо-6-фосфата. Если прокариоты выращивать на средах, где источник углерода — одно-, двух- или трёхуглеродные соединения, то необходимые сахара (в первую очередь С6) они должны синтезировать из имеющихся в среде источников углерода. У подавляющего большинства автотрофов на среде с двуокисью углерода (СО2) в качестве единственного источника углерода сахара синтезируются в реакциях восстановительного пентозофосфатного цикла. У гетеротрофов на среде с С2 и С3-соединениями для синтеза необходимых сахаров используются в значительной степени реакции катаболических процессов, например в гликолитическом пути. Слайд №21 Процесс, обеспечивающий синтез С6-углеводов из неуглеводных соединений, например, аминокислот, глицерина, молочной кислоты, получил название глюконеогенеза.
2. 5 Биосинтез жиров и липидов Слайд №22.Жирные кислоты с длинной цепью синтезируются путем конденсации и восстановления ацетатных групп. Для повышения реакционной способности метильная группа ацетилкофермента А сначала карбоксилируется в биотинзависимой реакции с образованием малонил-кофермент А:
+ НООС-СН2-СО ~ SCO А + ADP + Рi В последующих реакциях конденсации карбоксильная группа снова отщепляется в виде двуокиси углерода. Синтез жирных кислот происходит при участии мультиферментного комплекса.
Ацетил-СоА + 7 Малонил-СоА + 14NADPH2 -> Пальмитил-СоА + 14NADP + 7С02 + 7СоА + 7Н20 Жирные кислоты синтезируются отдельно, а затем с помощью эфирной связи включаются в липиды. Число типов жирных кислот у каждого вида бактерий строго определено. У прокариот преимущественно встречаются насыщенные жирные кислоты. Полиненасыщенные жирные кислоты, содержащие две и более двойных связей, найдены только у циакобактерий. Пути, ведущие к синтезу фосфолипидов, состоят из нескольких этапов. Исходным субстратом служит фосфодиоксиацетон (промежуточное соединение гликолитического пути), восстановление которого приводит к образованию 3-фосфоглиперина. К последнему затем присоединяются два остатка жирных кислот. Продуктом реакции является фосфатидная кислота. Активирование ее и последующее присоединение к фосфатной группе серина, инозита, глицерина или другого соединения приводят к синтезу фосфатидилсерина, фосфатидилинозита и фосфатидилглицерина соответственно. 3. Пути метаболизма, приводящие к образованию макроэргов 3.1 Энергетические ресурсы микробной клетки Микроорганизмы могут использовать не все виды энергии, существующей в природе. Недоступными для них являются ядерная, механическая, тепловая виды энергии. Доступными внешними источниками энергии являются электромагнитная энергия (свет определенной длины волны) (физическая) и химическая (восстановленные химические соединения). Способностью использовать энергию света обладает большая группа фотосинтезирующих организмов, в том числе и прокариот, имеющих фоторецепторные молекулы нескольких типов (хлорофиллы, каротиноиды, фикобилипротеины). Для всех остальных организмов источниками энергии служат процессы окисления химических соединений. Часто источниками энергии служат биополимеры, находящиеся в окружающей среде (полисахариды, белки, нуклеиновые кислоты), а также липиды. Прежде чем быть использованными, биополимеры должны быть гидролизованы до составляющих их мономерных единиц. Этот этап весьма важен по следующим причинам. Белки и нуклеиновые кислоты отличаются исключительным разнообразием. Количество видов белков исчисляется тысячами, после гидролиза, же образуется только 20 видов аминокислот. Все разнообразие нуклеиновых кислот (ДНК и РНК) после гидролиза сводится к 5 видам нуклеотидов. Таким образом, расщепление полимеров до мономерных единиц резко сокращает набор химических молекул, которые могут быть использованы организмом. 3.2 Общая характеристика энергетических процессов Слайд №23. В самом общем виде процессы, способные служить источником энергии для прокариот, можно представить следующим образом: А —> В + е-. Например, Fe2+ → Fe3+ + е--; (1) -СН2-СН2- → -СН=СН- + 2е--; (2) СН4 + 1/2 02 → СН3ОН. (3)
В первой реакции окисление иона двухвалентного железа — это потеря электрона. Во втором примере окисление углеродного субстрата можно в равной мере рассматривать как отрыв от него водорода (дегидрирование) или независимое удаление двух протонов (Н+) и электронов (е-). В биохимических процессах, как правило, перенос водорода осуществляется путем раздельного транспорта протонов и электронов: протоны выделяются в среду и при необходимости поглощаются из нее, электроны обязательно должны быть переданы на соответствующие молекулы. Поэтому все окислительно-восстановительные превращения определяются по существу " перемещениями" электронов. Реакции, в которых имеется возможность отрыва электронов, могут быть использованы прокариотами для получения энергии. Разнообразные соединения, способные окисляться, т.е. являющиеся источниками отрываемых электронов, называются донорами электронов. Поскольку электроны не могут существовать самостоятельно, они обязательно должны быть перенесены на молекулы, способные их воспринимать и, таким образом, восстанавливаться. Такие молекулы называются акцепторами электронов Слайд №24. У прокариот известны три способа получения энергии: разные виды брожения, дыхания и фотосинтеза. В процессах брожения в определенных окислительно-восстановительных реакциях образуются нестабильные молекулы, фосфатная группа которых содержит много свободной энергии. Эта группа с помощью соответствующего фермента переносится на молекулу АДФ, что приводит к образованию АТФ. Реакции, в которых энергия, освобождающаяся на определенных окислительных этапах брожения запасается в молекулах АТФ, получили название субстратного фосфорилирования. Их особенностью является катализирование растворимыми ферментами. Образующийся в восстановительной части окислительно-восстановительных преобразований сбраживаемого субстрата восстановитель (НАД.Н2, восстановленный ферредоксин) переносит электроны на подходящий эндогенный акцептор электрона (пируват, ацетальдегид, ацетон и др.) или освобождается в виде газообразного водорода (H2). Слайд №25. Многие прокариоты получают энергию в процессе дыхания. Они окисляют восстановленные вещества с относительно низким окислительно-восстановительным потенциалом (Е0), которые возникают в реакциях промежуточного метаболизма или являются исходными субстратами, например НАД.Н2, сукцинат, лактат, NH3, Н2S и др. (Таблица 1). Окислительно-восстановительный потенциал (Е0) характеризует способность определенных веществ быть донорами или акцепторами электронов. Он может быть измерен экспериментально для любой окислительно-восстановительной Для измерения окислительно-восстановительных потенциалов в качестве стандартного электрода обычно используют водородный электрод — раствор, содержащий ионы водорода (Н+) с активностью, равной 1 (рН=0), и электрод из инертного металла в равновесии с молекулярным водородом (Н2)при давлении 1атм. Его окислительно-восстановительный потенциал условно принят за 0 (Е0 = 0 В). Спонтанно или в присутствии соответствующих катализаторов окислительно-восстановительные системы с отрицательными окислительно-восстановительными потенциалами восстанавливают протон (Н) до молекулярного водорода (н2). Окислительно-восстановительные системы с положительным электрохимическим потенциалом сами восстанавливаются до молекулярного водорода (Н2). Окисление происходит в результате переноса электронов через локализованную в мембране дыхательную электронтранспортную цепь, состоящую из набора переносчиков, и приводит в большинстве случаев к восстановлению молекулярного кислорода до H20. Таким образом, в процессе дыхания молекулы одних веществ окисляются, других — восстанавливаются, т. е. окислительно-восстановительные процессы в этом случае всегда межмолекулярны. Наиболее широко распространена у прокариот способность окислять органические субстраты (гетеротрофы). Обнаружены также весьма специализированные группы прокариот, способные окислять различные неорганические субстраты (Н2, NH4+, NO2 H2S, S0, S2O32-, Fe2+ и др.) (автотрофы) с соответствующим восстановлением кислорода до Н2О. Наконец, прокариоты могут окислять органические и неорганические вещества с использованием в качестве конечного акцептора электронов не молекулярный кислород, а целый ряд органических и неорганических соединений (фумарат, СО2, NO3-, S°, SO42-, SО32- и др.). Количество освобождающейся энергии определяется градиентом окислительно-восстановительных потенциалов при переносе электронов от донора к акцептору. Так, окисление иона водорода (Н) молекулярным кислородом сопровождается освобождением значительно большего количества свободной энергии (∆ G0'= -238 кДж/моль), чем окисление никотинамиддинуклеотида (НАД-Н+) фумаратом (∆ G0'= -68 кДж/моль). Слайд №26. У прокариот известны три типа фотосинтеза: I — зависимый от бактериохлорофилла бескислородный фотосинтез, осуществляемый разными видами зеленых, пурпурных бактерий и гелиобактериями; II — зависимый от хлорофилла кислородный фотосинтез, свойственный цианобактериям и прохлорофитам; III - зависимый от бактериородопсина бескислородный фотосинтез, найденный у экстремально галофильных архебактерий. В основе фотосинтеза I и II типа лежит поглощение солнечной энергии различными пигментами, приводящее к разделению электрических зарядов, возникновению восстановителя с низким и окислителя с высоким окислительно-восстановительным потенциалом. Перенос электронов между этими двумя компонентами приводит к выделению свободной энергии. В фотосинтезе III типа окислительно-восстановительные переносчики отсутствуют. В этом случае энергия в доступной для организма форме возникает в результате светозависимого перемещения иона водорода через мембрану. Изучение у прокариот злектронтранспортных цепей, функционирующих в процессах дыхания и фотосинтеза I и II типа, выявило принципиальное сходство между ними. В обеих системах электронного транспорта есть флавопротеины, хиноны, цитохромы и белки, содержащие негемовое железо, позволяющие переносить электроны вниз по термодинамической лестнице. Таким образом, по существу электронтранспортные цепи 1 и 11 типов являются окислительными. Разнообразие в их организации обнаружено в широком наборе доноров и акцепторов электронов, и в конкретной организации самих цепей: химическом строении переносчиков, принадлежащих к одному типу, их наборе, расположении ит.д. В процессах дыхания и фотосинтеза освобождающаяся при переносе электронов энергия запасается первоначально в форме электрохимического трансмембранного градиента ионов водорода (∆ μ н+ ) т. е. имеет место превращение химической и электромагнитной энергии в электрохимическую. Последняя затем может быть использована для синтеза АТФ. Поскольку в обоих процессах синтез АТФ обязательно связан с мембранами, реакции, приводящие к его образованию, получили название мембранзависимого фосфорилирования. Последнее подразделяется на два вида: окислительное фосфорилирование (АТФ образуется в процессе электронного переноса при окислении химических соединений) и фотосинтетическое фосфорилирование (синтез АТФ связан с фотосинтетическим электронным транспортом). Следует подчеркнуть, что принципы генерации АТФ при фотосинтезе и дыхании, т.е. механизмы мембранзависимого фосфорилирования, одинаковы. Таким образом, энергия, получаемая в процессах брожения, дыхания или фотосинтеза, запасается в определенных формах. Слайд №27. Существуют две универсальные формы энергии, которые могут быть использованы в клетке для выполнения разного рода работы: энергия высокоэнергетических химических соединений (химическая) и энергия трансмембранного потенциала ионов водорода (электрохимическая).
|
Последнее изменение этой страницы: 2017-04-12; Просмотров: 567; Нарушение авторского права страницы