|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Рецепторы стероидных гормонов: регуляция транскрипции
Рецепторы стероидных гормонов хорошо демонстрируют, как развивается и меняется наши представления о регуляции транскрипции у эукариот. До примерно конца 80-х прочно держалось представление о том, что гормоны, связанные с рецептором, только и делают, что активируют транскрипцию. Оказалось, что это не так. Первыми генами, для которых было показано, что они негативно регулируются глюкокортикоидными гормонами были гены проопиомеланокортина, альфа-субъединицы гликопротеиновых гормонов и гена коллагеназы типа I. Таким образом одни и теже регуляторы могут и активировать, и репрессировать транскрипцию в зависимости от конкретного окружения, в которых регулятор оказывается в клетке. В целом выделяют три стратегии, которые используют рецепторы стероидных гормонов для регуляции активности генов: 1.Активация транскрипции генов путем связывания рецептора, содержащего лиганд, с позитивно регулируемым GRE. 2.Репрессия транскрипции за счет связывания с негативно-регулируемым GRE. 3.Регуляция за счет взаимодействия рецепторов не с GRE, а с участками связывания других транскрипционных факторов. Если стероидные рецепторы образуют гомодимеры, которые участвуют в активации транскрипции, то тироидные гормоны, ретиноевая кислота и витамин В работают посредством рецепторов, образующих гетеродимеры. Но в целом все эти рецепторы образуют суперсемейство лиганд зависимых регуляторов транскрипции. Тироидные рецепторы также не только активируют, но могут и репрессировать транскрипцию. Это они делают в отсутствии лиганда.
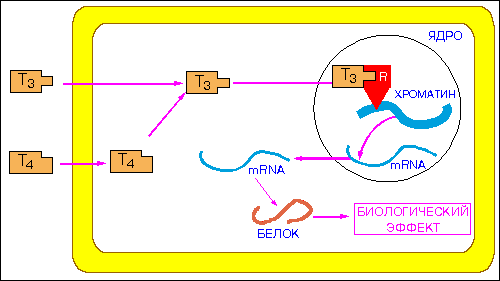
Как же происходит индукция генов под действием стероидных и тироидных гормонов? Лучше всего это изучено для глюкокортикоидного рецептора. Появляясь в окружении клетки, гормон проникает в нее путем простой диффузии. Внутри клетки он связывается с рецептором, и транспортируется в клеточное ядро рис. стероидный гормон проникает в ядро.
Рецепторы связываются со специфическими последовательностями в промоторах или энхансерах. В результате активируется (или репрессируется) экспрессия тех генов, которые должны реагировать в ответ на действие стероидного гормона. Обычно его элемент отклика GREнаходится в энхансерах. Это несовершенный палиндром (см Табл 2 svtr ). С ним связывается димер рецептора в комплексе с лигандом. Точнее сказать димеризация лиганд-связанного рецептора происходит при связывании с палиндромной последовательностью GRE [Karin ea 1998]. При связывании происходит активация энхансера. *-*-*- энхансеры -протяженные последовательности нуклеотидов, которые содержат сайты связывания нескольких факторов транскрипции. Характерными свойствами энхансера являются его способность осуществлять регуляторное действие на промотор на больших расстояниях от него (более 60 т.п.о.), независимость его активности от ориентации по отношению к промоторам и от расположения относительно регулируемого гена. На рис. I.30
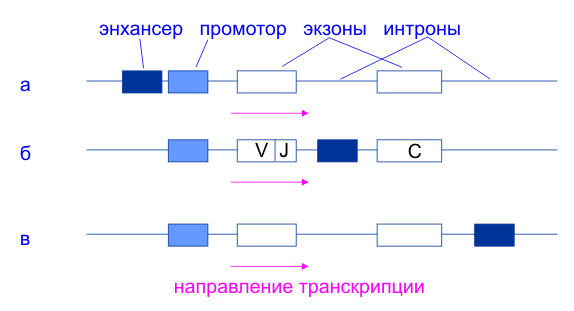
приведены схемы строения нескольких эукариотических генов, отражающие взаимное расположение структурных и регуляторных частей. У гена альбумина ( рис. I.30, а ) энхансер находится перед промотором, а регуляторная часть предшествует его структурной части. У генов иммуноглобулинов регуляторные элементы локализованы в интронах самого гена ( рис. I.30, б ). Энхансер может быть расположен и ниже гена на значительном от него расстоянии, как у бета-глобинового гена ( рис. I.30, в ). Энхансеры являются своеобразными матрицами для сборки сложных белковых комплексов, структура которых обеспечивает высокоспецифические белок-белковые связывания и передачу регуляторных сигналов РНК-полимеразе II , находящейся в составе инициационного комплекса. Энхансер ( Grosschedl ea, 1980 ) это генетический цис-элемент, обладающий усиливающим транскрипцию действием, которое практически не зависит от расположения элемента относительно контролируемого им гена. Энхансеры представлены короткими последовательностями ДНК, состоящими из отдельных элементов (модулей), включающих десятки нуклеотидных пар. Модули могут представлять собой повторяющиеся единицы ( Herr ea, 1986 ). Энхансер увеличивает эффективность транскрипции гена в десятки и сотни раз. Особенность энхансеров состоит в том, что они способны действовать на больших расстояниях (более чем 1000 п.н.) и вне зависимости от ориентации по отношению к направлению транскрипции гена. Энхансеры могут располагаться как на 5'-, так и на 3'-конце фрагмента ДНК, включающего ген, а также в составе интронов. Энхансеры обнаружены на 3'-фланге гена, кодирующего полипептидный гормон - плацентарный лактоген человека, а также, в составе интронов генов иммуноглобулинов и коллагена. Энхансеры - основное средство регуляции транскрипции ( Gluzman ea, 1983; Khoury ea, 1983 ) в клетках высших эукариот. Энхансеры были первоначально определены, как цис-действующие элементы, которые усиливают транскрипцию независимо от их ориентации и расстояния от гена. Например энхансер " wing margin" cut локуса Дрозофилы находится на расстоянии 85 кб перед промотором, тогда как энхансер мышиного иммуноглобулинового гена Hмю находится в одном из интронов гена. Энхансер альфа цепи Т-клеточного рецептора находится на расстоянии 69 кб в 3'-сторону от промотора. Более того, спаривание хромосом позволяет иногда активному энхансеру на одной хромосоме активировать транскрипцию аллельного промотора на другой[ Blackwood ea 1998 ]. Еще одной важной характеристикой энхансера является его способность активировать любой промотор, расположенный сравнительно недалеко. В целом можно сказать, что энхансер действует на ближайший к нему промотор. Эффект регуляции достигается тем же путем, что и регуляция проксимальными элементами промоторов, т.е. путем сборки на последовательностях энхансера белкового комплекса, который иногда называют энхансеосомой [ Werner ea 1997 ], и его взаимодействия с основным транскрипционным комплексом путем белок-белковых взаимодействий. Ясно, что для того чтобы осуществить эти контакты ДНК, должна изогнуться. Исходно существовало представление, что энхансеры действуют вне зависимости от ориентации относительно промоторов. В последнее время появились данные, показывающие, что существует стереоспецифичность во взаимодействии энхансерных белковых комплексов, и что благодаря этой стереоспецифичности небезразлично, как направлены многие энхансеры по отношению к промотору, который они регулируют. Неясно, насколько эффект стереоспецифичности распространен, но сам факт ее существования, хотя бы в ряде случаев, говорит о том, что основная концепция независимости эффекта энхансера от ориентации не является универсальной. Контрольные области генов ( LCR ) часто содержат множественные автономные энхансерные модули которые варьируют в размерах от 50 по до 1, 5 кб [ Blackwood ea 1998 ]. Каждый из этих модулей предназначен для выполнения определенных функций. Он может активировать ген на определнной стадии развития или в определенном типе клеток. Отдельный ген может таким образом содержать множество энхансерных модулей, каждый из которых вносит свой вклад в пространсвенную и временную регуляцию экспрессии гена. Вслед за авторами обзора [Blackwood ea 1998 ] здесь под энхансером я буду подразумевать отдельный модуль. Часто энхансеры содержат те же модули, что и промоторы. *** У вируса простого герпеса (ВПГ) обнаружен транскрипционный фактор VP16, участвующий в активации его непосредственно-ранних генов и присутствующий в зрелом вирионе в количестве около 1000 копий. Белок VP16 взаимодействует с энхансерами, расположенными перед каждым из непосредственно-ранних генов вируса, что приводит к активации их транскрипции. Каждый из энхансеров содержит одну или несколько копий цис-регуляторных последовательностей - нонануклеотидную последовательность 5'-TAATGARAT-3' ( " tat-garat" ) и прямые повторы гексануклеотидной последовательности 5'-CGGAAR-3' ( " cigar" ). В очищенном состоянии белок VP16 не взаимодействует ни с одной из них. В присутствии белковых факторов экстрактов ядер клеток-хозяев VP16 входит в состав многокомпонентного белкового комплекса, формирующегося на последовательности " tat-garat" ( рис. I.31 ). Сходство последовательностей нуклеотидов мотива " tat-garat" и сайтов связывания Oct-белков, обнаруженных в регуляторных последовательностях генов гистонов и иммуноглобулинов, заставило проверить эти белки на способность взаимодействовать с ВПГ-энхансерами. Оказалось, что Oct-1 является вторым белковым компонентом, соединяющимся с " tat- garat" -последовательностью энхансера и облегчающим связывание с ним белка VP16.
Эффективное образование белкового комплекса требует участия других компонентов клетки-хозяина, поскольку очищенные белки Oct- 1, VP16 и последовательность " tat-garat" формируют стабильный комплекс только в присутствии экстрактов ядер клеток HeLa. Белковый фактор, названный HCF, может непосредственно взаимодействовать с VP16 в отсутствие других компонентов комплекса. С помощью мутационного анализа установлено, что 5'- концевой сегмент " tat-garat" ( последовательность " tat" ) необходим для связывания белка Oct-1, тогда как ее 3'-концевая последовательность ( " garat" ) требуется для образования полного белкового комплекса Oct-1-VP16-HCF. Таким образом, четыре молекулярных поверхности (интерфейса) участвуют в формировании функционально активного комплекса на матричной поверхности мотива " tat-garat" энхансеров непосредственно-ранних генов ВПГ. Одна группа контактов образуется между белками Oct-1 и VP16, вторая - между VP16 и HCF, третья - между Oct-1 и последовательностью " tat" и четвертая - между VP16 и последовательностью " garat" ( рис. I.31 ).***
Сложная серия молекулярных интерфейсов участвует в формировании функционального макромолекулярного комплекса на мотиве " cigar" энхансера непосредственно-ранних генов ВПГ. Белок, взаимодействующий с этим мотивом, выделен из экстрактов ядер печени крыс и получил название GABP (GA-binding protein). Он состоит из двух субъединиц - GABPальфа и GABPбета. Белковые комплексы, похожие на GABP, выделены и из культивируемых клеток человека. Часть аминокислотной последовательности GABPальфа оказалась гомологичной участку ДНК-связывающего домена транскрипционных факторов, принадлежащих к семейству ETS. Все представители этого семейства обладают последовательностью из 85 аминокислотных остатков, которая необходима для их специфического взаимодействия с ДНК. Таким образом, ETS-домен белка GABPальфа образует первый молекулярный интерфейс, обеспечивающий связывание белка с мотивом " cigar" энхансера. В смеси с GABPбета альфа-субъединица образует белковый комплекс, который обладает бoльшим сродством к мотиву " cigar" энхансера, чем сама альфа-субъединица. Анализ особенностей взаимодействия GABP-комплекса с энхансером позволил обнаружить три высокоспецифичных молекулярных интерфейса: домен полипептида GABPальфа, необходимый для его взаимодействия с белком GABPбета и включающий ETS-домен, и последовательность из 35 С-концевых аминокислотных остатков GAPBальфа. У полипептидной цепи GABPбета во взаимодействии с GABPальфа принимают участие четыре тандемных повтора, состоящие из 33 аминокислотных остатков каждый и расположенные в N-концевой части белка. Анализ аминокислотных последовательностей, гомологичных этому повтору у ряда других белков, позволил выявить высококонсервативную последовательность TPLH. Дополнительные молекулярные взаимодействия между белковым комплексом и последовательностью нуклеотидов " cigar" осуществляются через повторы TPLH после того, как они образуют требуемый комплекс с GABPальфа. При этом аминокислотные последовательности повторов непосредственно контактируют с ДНК. Наконец, в последнем из известных контактов, имеющих место после сборки белкового комплекса на последовательности нуклеотидов " cigar", участвует так называемый домен димеризации полипептидной цепи GABPбета, расположенный вблизи ее С-конца. Очищенные субъединицы GABPбета в растворе ассоциируют с образованием гомодимеров, которые в присутствии субъединиц GABPальфа формируют гетеродимерный комплекс альфа2бета2, хорошо приспособленный для взаимодействия с гексануклеотидным повтором " cigar" (CGGAAR)2 энхансера. Анализируя все вышеперечисленные компоненты энхансеров непосредственно-ранних генов ВПГ, можно сделать вывод, что в их активации принимают участие не менее пяти белков (VP16, Oct-1, HCF, GABPальфа и GABPбета), которые участвуют, как минимум, в пяти белок-белковых контактах и шести специфических контактах белок-ДНК ( рис. I.31 ). Прежде всего, обращает на себя внимание кооперативность взаимодействия компонентов, активирующих энхансеры. Ключевые белки этого комплекса - Oct-1, VP-16 и GABPальфа сами по себе слабо взаимодействуют со специфическими последовательностями ДНК, но обладают способностью собираться в стабильный функционально активный комплекс двумя путями: во- первых, объединяясь с кофакторами, которые самостоятельно не способны взаимодействовать с ДНК (например, VP16 с HCF и GABPальфа с GABPбета); во-вторых, контактируя с последовательностью нуклеотидов энхансера, содержащей функциональные участки " tat" и " garat", а также повтор " cigar" и образующей матрицу для геометрически правильной сборки всего активирующего комплекса. -*-*-* Основные механизмы действия энхансеров: считается, что функциональные участки генома, содержащие один или несколько генов, образуют длинныепетли, включающие десятки тысяч нуклеотидных пар ДНК. Высказано представление, что петли закреплены в матриксе клеточного ядра и могут быть сверхспирализованы. В состав матрикса входит топоизомераза II, по-видимому, определяющая топологию петли ДНК. В таком случае взаимодействие энхансера с белками может менять конформацию всей петли, включая и удаленный от энхансера участок ДНК, в результате чего в составе петли изменяется локальная структура хроматина и облегчается транскрипция гена. Более вероятно, что влияние энхансера, связанного с белком, определяется его непосредственным взаимодействием с РНК-полимеразой и другими факторами транскрипции в процессе инициации. Такое взаимодействие может осуществляться благодаря сгибанию молекулы ДНК, что создает возможность непосредственного контакта районов промотора и удаленного от него энхансера, связанных со специфическими белками. По-видимому множество различных обстоятельств определяют активность энхансеров. Взаимодействия факторов с последовательностями энхансеров. С факторами, связывающимися с ДНК, в свою очередь, могут взаимодействовать множество ко-активаторов, которые прямо не взаимодействуют с ДНК. По-видимому существуют прямые белок - белковые контакты между факторами, связанными с энхансерами и компонентами базального транскрипционного комплекса. Такие контакты возможны благодаря сгибанию ДНК. Эти взаимодействия, вероятно, приводят к образованию больших транскрипционных комплексов, с которых транскрипция легко инициируется и реинициируется. Энхансеры могут влиять на ковалентную модификацию белков. Мы видели, например, что С-концевой домен РНК-полимеразы II интенсивно фосфорилируется на ранних стадиях транскрипции. Ацетилирование гистонов уменьшает репрессирующую роль хроматина в транскрипции. Многие активаторы и ко-активаторы транскрипции обладают гистон ацетилтрансферазной активностью, тогда как некоторые ко-репрессоры проявляют гистон деацетилазную активность. Гистоны не единственный элемент, который ацетилируется и деацетилируется. Ацетилируется например р53 антионкоген, а также TFIIE и TFIIF факторы базальной транскрипции. Белковые факторы, связывающиеся с энхансерами, могли бы потенциально влиять на эти процессы. Энхансеры могут влиять на структуру хроматина , которая может обеспечивать взаимодействия факторов на больших расстояниях благодаря компактизации ДНК. Ремоделирование нуклеосом также может быть одним из механизмов проявления энхансерной активности. Хроматин ремоделирующие факторы известны: SWI-SNF, NURF, RSC, ACF и CHRAC. Некоторые из этих факторов могут действовать глобально в масштабах генома, тогда как другие активны только в окружении данного гена. Еще один механизм может заключаться в том, что энхансеры могут выставлять подконтрольные им промоторы в такие домены ядра, которые содержат высокие локальные концентрации транскрипционных факторов. Способность ряда энхансеров взаимодействовать со специфическими белками дифференцированной клетки, вероятно, обеспечивает их важное свойство - тканевую специфичность.
Индукция транскрипции В ответ на внешние сигналы включаются (или выключаются) отдельные гены или группы генов. Эти группы генов таким образом находятся под общим контролем. Это означает, что у всех них есть общие элементы, узнаваемые транскрипционными факторами, которые появляются или исчезают в ответ на индуцирующий сигнал извне. Такие ДНК участки называют элементами отклика - response elements, RE. Это могут быть элементы отклика на тепловой шок, на стероидные или тироидные гормоны, на интерфероны, на тяжелые металлы и т.д. Элементы отклика могут находиться в промоторе или энхансере. Они содержат довольно короткие последовательности, сходные для элементов одного типа.
ко-активатор стероидных рецепторов ( SRC, steroid-receptor co-activator ), который непосредственно савязываются с AF-2 областью нескольких ядерных рецепторов, связанных со своими лигандами. Этот ко-активатор конститутивно ассоциирован с CREB связывающим белком, CREB-binding protein, CBP.
В промоторной области ряда сАМР-зависимых генов были обнаружены так называемые CRE (сАМР regulatory elements) - участки, регулируемые cAMP и обладающие свойствами энхансеров с консенсусом TGACGTCA
Ответ гена c-fos на агенты, повышающие уровень cAMP в клетке, опосредуется в основном элементами CRE (цАМФ регулирующий элемент) в промоторной области гена. С элементом CRE в позиции -64/-57 связываются CRE-связывающие белки CREB.
-*-*-*-CREB-белки кодируются как минимум десятью различными генами, и это семейство включает помимо CREB так называемые активирующие транскрипционные факторы ( ATF ) [ Foulkes N.S. ea, 1994 ]. CREB-Белки, подобно Fos- и Jun- белкам, имеют мотив " лейциновой молнии" ( ZIP ), необходимый для димеризации [ Abel T. and Maniatis T., 1989 ], и связываются с CRE в димерной форме [Foulkes N.S. ea, 1994 ]. Примечательно, что они могут также димеризоваться с белком c-Jun. Jun/CREB гетеродимер обладает большим сродством к CRE, чем к TRE [ Macgregor P.F. ea, 1990 ]. Белок CREB содержит домен, названный KID (" kinase-inducible domain" ) или Р-бокс, необходимый для его РКА (Протеинкиназа А) - зависимой активации, в котором расположен Ser-133 - сайт для фосфорилирования PKA (протеинкиназа А, цАМФ зависимый) [ Lalli E. and Sassone-Corsi P., 1994, Brindle P. ea, 1994 ].-*-*-* Существуют несколько членов семейства CRE-связывающих белков, ингибирующих транскрипцию и названных " CRE- модуляторами" ( CREM ). Наименьшие размеры среди всех CRE-связывающих белков имеет cAMP- индуцибельный ранний репрессор ( ICER ), содержащий только домены ДНК- связывания и димеризации и также определяющий подавление транскрипции [ Lalli E. and Sassone-Corsi P., 1994, Molina C.A. ea, 1993 ]. CREB, белки могут фосфорилироваться и PKC. PKC (протеинкиназа С) может фосфорилировать CREB по Ser-133, а также аналогичный сайт в CREM-тау [Lalli E. and Sassone-Corsi P., 1994]. РКС может фосфорилировать также и другие CRE-связывающие факторы, в частности, CRE-BP1 [ Sakurai A. ea, 1991 ]. Таким образом, возможно, РКС может участвовать в регуляции активности гена c-fos, фосфорилируя CRE-связывающие трансактиваторы, хотя необходимо отметить, что данная регуляция в физиологических условиях не показана. Однако, существует большое количество акцепторов кальция помимо РКС, в частности, кальмодулин, регулирующий протеинкиназы, относящиеся к семейству кальмодулинзависимых протеинкиназ ( СаМК ) [ Pietrobon D. ea, 1990 ]. Показано, что деполяризация клеток РС-12 приводит к повышению активности CREB, и СаМК фосфорилируют этот транскрипционный фактор по Ser- 133 in vitro [Sheng M. ea, 1991]. Однако изоформа СаМКII, по-видимому, не играет большой роли в стимуляции CREB, поскольку эта изоформа не обнаруживается в клеточном ядре и, кроме того, фосфорилирует CREB также и по Ser-142, что блокирует его активацию. В то же время СаМКIV, способная мигрировать в ядро, фосфорилирует CREB только по Ser-133, способствуя повышению его активности [Matthews R.P. ea, 1994, Sun P. ea, 1994 ]. Кроме РКА и СаМК обнаружена протеинкиназа, которая фосфорилирует CREB по Ser-133 в ответ на NGF и EGF в клетках РС-12. Эта протеинкиназа в отличие от перечисленных не является непосредственным эффектором вторичных мессенджеров. Она функционирует как компонент Ras-зависимого пути передачи сигнала и отлична также и от RSK и pp70(S6K) [ Ginty D.D. ea, 1994 ]. Таким образом, cAMP-чувствительные элементы в РС-12 клетках могут контролироваться без участия РКА и СаМК. -*-*- |
Последнее изменение этой страницы: 2017-04-13; Просмотров: 660; Нарушение авторского права страницы