|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Особенности биокаталитических процессов в живых организмах. Принципы структурной организации ферментов. Специфичность, стереоспецифичность. Фермент-субстратный комплекс. Энергия активации.
Реакции, протекающие в живых организмах, подчиняются общим законам химии, однако отличаются высокой специфичностью и отсутствием побочных продуктов. Эта особенность определяется свойствами белковых посредников биохимических реакций – ферментов, выполняющих роль катализаторов. Ферменты представляют собой природные биокатализаторы. В настоящее время в биологических системах обнаружено несколько тысяч индивидуальных ферментов, 3709 из них – выделено, изучено и внесено в специальную базу данных. Подсчитано, что живая клетка может одновременно содержать до 1000 различных ферментов, каждый из которых ускоряет ту или иную химическую реакцию. В химическом плане большинство ферментов имеют белковую природу, однако из этого правила есть исключение – рибозимы. Рибозимы – низкомолекулярные молекулы РНК, способные осуществлять низкотемпературный катализ. Ферменты как биокатализаторы имеют черты сходства и различия с неорганическими химическими катализаторами. Простые ферменты — это простые белки, они построены из аминокислот и при гидролизе распадаются только на аминокислоты. Сложные ферменты — это сложные белки, они состоят из простого белка и небелкового компонента. При их гидролизе, помимо свободных аминокислот, освобождается небелковая часть или продукты её распада. Белковая часть сложного фермента получила название апофермент, небелковая часть — кофактор. Кофакторы могут иметь разную химическую природу и отличаться по прочности связи с апоферментом. В роли кофактора могут выступать ионы различных металлов, а также другие неорганические ионы. Органические вещества неаминокислотной природы, используемые в роли кофакторов, называются коферментами. Кофермент вместе с апоферментом образуют холофермент. Уникальные свойства ферментов (высокая каталитическая активность и субстратная специфичность) обусловлены сложным пространственным строением и наличием в составе молекул определенных комбинаций химических групп. В структуре белков можно условно выделить несколько уровней организации.
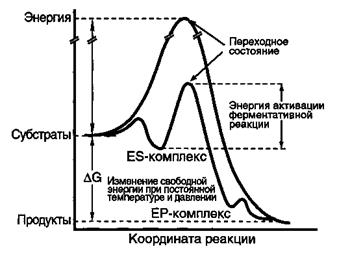
Специфичность ферментов: субстратная; действия. Субстратная специфичность — способность фермента катализировать превращения только одного определенного субстрата или же группы сходных по строению субстратов. Определяется структурой адсорбционного участка активного центра фермента. Типы субстратной специфичности: - абсолютная — способность фермента катализировать превращение только одного, строго определенного субстрата; - относительная — способность фермента катализировать превращения нескольких сходных по строению субстратов; - стереоспецифичность — способность фермента катализировать превращения определенных стереоизомеров (Фермент амилаза действует только на α -гликозидные связи, что позволяет гидролизовать крахмал и гликоген (полимеры глюкозы), остатки глюкозы в которых соединены α -гликозидными связями. Целлюлоза - также полимер глюкозы, однако остатки глюкозы в нём связаны β -гликозидными связями. В результате отсутствия у человека ферментов, специфичных к β -гликозид-ной связи, целлюлоза не гидролизуется в кишечнике человека и не может служить источником глюкозы). Специфичность действия — способность фермента катализировать только определенный тип химической реакции. В соответствии со специфичностью действия все ферменты делятся на 6 классов. Классы ферментов обозначаются латинскими цифрами. Название каждого класса ферментов соответствует этой цифре. Образование фермент-субстратного комплекса происходит в результате ряда конформационных изменений в структуре белка. Достигается за счет множественных контактов между ферментом и субстратом, причем, чем больше этих контактов, тем эффективнее протекает каталитический процесс. Связывание субстрата в активном центре фермента за счет слабых нековалентных взаимодействий и сопровождается ↓ свободной энергии. Конформация фермента существенно определяет его биологическую активность. Многочисленные экспериментальные факты свидетельствуют о том, что фермент способен выполнять свойственную ему каталитическую функцию, только находясь в определенной трехмерной структуре. Этот принцип, в частности, лег в основу двух моделей фермент-субстратного взаимодействия: так называемой модели «замка и ключа», предложенной Фишером, и общепринятой в настоящее время теории индуцированного соответствия, выдвинутой в середине прошлого века Кошландом. Модель Фишера достаточно наглядно объясняет высокую специфичность ферментов. Согласно этой модели, фермент обладает специфическим участком – «замком», который по своей структуре комплементарен молекуле субстрата – «ключу». Следовательно, с ферментом могут связываться и подвергаться дальнейшим каталитическим превращениям только субстраты, обладающие необходимой пространственной структурой. Если фермент находится в условиях, отличающихся от его обычного окружения, он может подвергаться структурным изменениям, которые обычно сопровождаются потерей его каталитической активности. В теории Фишера активный центр фермента рассматривается как жесткая структура, строго комплементарная структуре субстрата. Согласно теории Кошланда, исходная структура активного центра фермента не соответствует строго субстрату. Однако присутствие субстрата индуцирует структурные изменения активного центра, обеспечивающие связывание субстрата. Одновременно с этим происходит изменение структуры субстрата, благодаря чему достигается его комплементарность измененному активному центру. Предполагается, что активные центры большинства ферментов расположены в области, которая является более гибкой и мобильной, чем остальная часть молекулы белка. Результаты рентгеноструктурных исследований показали, что активные центры обычно располагаются в шарнирных областях, связывающих два структурных домена. В связи с этим активный центр фермента по сравнению со всей молекулой белка более чувствителен к действию денатурирующих агентов и физических факторов. Поэтому фермент может инактивироваться при относительно невысоких концентрациях денатурирующих агентов или при незначительных физических воздействиях, когда глобальные изменения конформации белковой молекулы еще не наблюдаются. Связывание субстрата в активном центре фермента обеспечивается слабыми нековалентными взаимодействиями (водородными связями, электростатическими взаимодействиями, вандерваальсовыми взаимодействиями и гидрофобными связями) и сопровождается уменьшением свободной энергии системы (Δ Gсвяз = –RTlnKравн). Для превращения субстрата в продукт следует затратить энергию, необходимую для ориентации реагирующих групп, образования нестабильных зарядов, перегруппировки связей и т. д. Это иллюстрируется «холмом» на энергетическом профиле реакции.
Участники реакции должны преодолеть этот барьер и перейти на более высокий энергетический уровень, то есть оказаться в переходном состоянии. Энергия, необходимая для достижения переходного состояния, называется энергией активации (Δ G±). Переходное состояние – это геометрическая и электронная структура, отражающая комбинированное состояние фермента и субстрата, в котором реализуется оптимальная ориентация реагирующих групп и происходит постоянное образование и разрыв связей. Концентрация переходного состояния и его время жизни крайне малы, в связи с чем переходное состояние трудно зафиксировать. Скорость реакции находится в обратной зависимости от величины энергии активации: чем выше Δ G±, тем ниже скорость. Это понятно, поскольку скорость реакции пропорциональна концентрации активированного комплекса, которая тем выше, чем ниже энергия активации. Мало с чем сравнимая эффективность ферментов как катализаторов объясняется их способностью снижать энергию активации и смещать равновесие в сторону образования переходного состояния. Достигается это благодаря очень прочному связыванию структуры переходного состояния. Энергия связывания переходного состояния (Δ Gсвяз = -RTlnKравн) является отрицательной величиной, поэтому ее возрастание будет снижать энергию активации: (Δ G± = Δ G±кат + (–RTlnKравн). |
Последнее изменение этой страницы: 2017-04-13; Просмотров: 1322; Нарушение авторского права страницы