|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Пространственная организация и функционирование фотосистем высших растений.
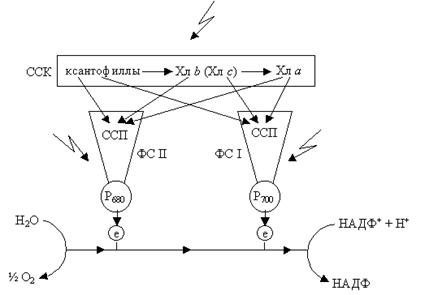
Пигменты в хлоропластах образуют у высших растений две системы. Каждая пигментная система составляет фотосинтетическую единицу, которая входит в фотосистемы I и II. Каждая из этих единиц состоит из набора молекул вспомогательных пигментов, передающих энергию на одну молекулу основного пигмента (Р700 и Р680). Последние молекулы входят в состав реакционных центров. Пигменты, входящие в состав систем, делят на пигменты-ловушки и пигменты-сборщики. Пигмент-ловушка может преобразовывать энергию. Пигмент-сборщик квантов света передает поглощенную энергию пигменту-ловушке. В пигментных системах ловушками являются Р700 и Р680. Остальные пигменты – сборщики. Перенос энергии возбуждения происходит в направлении от вспомогательных пигментов к хлорофиллу а, а затем к специальному хлорофиллу (Р700 и Р680), где происходят фотохимические реакции. Следовательно, общим результатом стадии является сбор энергии возбуждения, вызванной светом, и передача ее на реакционный центр. Наиболее общепризнанным механизмом передачи электронного возбуждения между молекулами пигментов является так называемый резонансный перенос. Для того чтобы произошел этот перенос энергии, возбужденная молекула должна индуцировать возбужденное состояние в другой молекуле, находящейся в непосредственной близости от нее. Перенос электронного возбуждения происходит тогда, когда индуцируется колебание электрона в другой молекуле. Когда перенос возбуждения завершен, колебания электрона в первой молекуле прекращаются и возникают колебания электрона в другой молекуле, которая в свою очередь переходит в возбужденное состояние. Надо же отметить, что для резонансного переноса электронного возбуждения необходимо, чтобы энергия, которую может передать возбужденная молекула, соответствовала энергии, которую может принять другая молекула. Две пигментные фотосистемы – это две машины, движущие фотосинтез на световой стадии. Каждая состоит из реакционного центра и ансамбля молекул – светособирающих пигментов (пигмент-белковый комплекс – ПБК. Каждой фотосистеме соответствует свой отдельный набор пигментов, а также связанных с ними переносчиков электронов и в каждой фотосистеме происходят свои, только ей присущие фотосинтетические реакции. В состав ФС I входят первая пигментная система с Р700, мономерная форма Хл а695, белки, содержащие железо и серу, 1–2 молекулы ферродоксина, по 1 молекуле цитохрома и пластицианина и 2 молекулы цитохрома b6; В состав ФС II – вторая пигментная система с Р680, 4 молекулы пластацианина, 2 молекулы цитохрома b559 и 6 атомов марганца. В ЭТЦ входит и пул пластахинонов – липидрастворимых переносчиков е– и Н+. В состав светособирающих пигментов (светособирающие антены) входят: каротин, Хл а662, а670, а677, а684, а692 (ФС I) и ФС II: а622, а670, а677. Кроме того, имеется еще светособирающий белковый комплекс (ССК) Считают, что при низкой интенсивности света собственные антенны ФС I и ФС II поглощают мало квантов света и поэтому не могут поддерживать необходимой скорости фотосинтезирующих реакций, и нужен дополнительный приток энергии, который обеспечит ССК. В состав ССК входят ксантофиллы, хлорофилл b, некоторое количество хлорофилла а. ССК передает поглощенную энергию на ПБК и затем на реакционный центр (рис. 2.10). Получив эту энергию, специальная форма хлорофилла в реакционном центре переходит в возбужденное состояние (Хл*). Хл* обладает очень высокой реакционной способностью и является сильным восстановителем.
В окислительно-восстановительной системе А/А– происходит восстановление А до А–. От А– электрон переходит дальше и через другие компоненты электрон-транспортной цепи в конечном счете переносятся на НАДФ+. Хлорофилл, который отдал электрон, переходит в состояние свободного катиона – радикала Х+. Он возвращается в основное первичное состояние, получив электрон восстановленной формы Д– – второй окислительно-восстановительной системы Д/Д–. Окисленный Д восстанавливается до Д–, получая электрон от воды ФС (II) и ФС (I) размещаются последовательно в цепи транспорта электронов от Н2О до НАДФ+. Для восстановления одной молекулы НАДФ+ в процессе фотосинтеза необходимо два электрона и два протона, при этом донором электронов является вода. Фотоиндуцированное окисление воды происходит в ФС II, восстановление НАДФ+ – ФС I. Таким образом, фотосистемы должны функционировать во взаимодействии. Исходя из данных квантового выхода фотосинтеза (количество выделенного О2 или связанного СО2 на 1 квант поглощенной энергии), необходимо восемь квантов света для выделения одной молекулы О2; из состава компонентов ФС I и ФС II и величины окислительно-восстановительного потенциала была разработана схема последовательности реакций в световой стадии фотосинтеза. Из-за подобия с буквой Z эта схема получила название Z-схемы Таким образом, фотохимическую работу у высших растений, выполняемую при фотосинтезе, в конечном счете, можно свести до разложения воды. Реакционный центр ФС II включает кислород выделяющий комплекс, хлорофилл Р680, первичный акцептор электронов феофитин (Фео) – производное Хл, в котором Mg замещен протонами, и вторичные акцепторы – молекулы монохинона (Qн и Qв), а также пластохинон (Q). Ядро реакционного центра ФС II составляют два мембранных белка (D1 и D2) с молекулярной массой (ММ) 32 и 34 кД. Оба белка имеют по пять трансмембранных петель и служат основой для связывания большинства простетических групп, выполняющих функции переносчиков электронов. Другие белки (43 и 47 кД) ФС II входят в состав комплекса светособирающих пигментов или участвуют в выделении кислорода при фотоокислении воды (33, 23, 16 кД). Функции остальных белков с небольшой ММ, например цитохрома b559 и других, пока неизвестны. В кислородвыделяющий комплекс входит Mn-содержащий пул и как кофакторы кальций и хлор. Посредником между кислородвыделяющим центром и хлорофиллом Р680 является остаток аминокислоты тирозина (Yz) белка D1 ( см. рис. 2.13). Реакционный центр ФС I включает первичный донор электронов Хл Р700, первичный акцептор – Хл а (А0), промежуточный акцептор – филлохинон (А1), вторичные акцепторы – железосерные белки (FX, FA, FB), водорастворимы железосерный белок ферредоксин (ФД), растворимый флавопротеин – ферредоксин-НАДФ-редуктазу (ФП). ФС I представляет собой интегральный пигментбелковый комплекс с ММ около 340 кД. Комплекс цитохром b6/f состоит из цитохрома b6, цитохрома f, железосерного белка Риске и субъединицы IV. Цитохром b6 и субъединица IV являются наиболее гидрофобными элементами комплекса. Простетическая группа цитохрома f представлена гемом с-типа, а аналогичная группа цитохрома b6 состоит из двух гемов b-типа, ковалентно связанных с остатками гистидина. Один гем называют высокопотенциальным (Н), второй низкопотенциальный (L). В передаче электронов от ФС II, а именно от комплекса b6/f к ФС I участвует пластоцианин (водорастворимый белок, содержащий два атома меди. Во время перемещения электрона по такой цепи переносчиков часть его энергии преобразуется в химическую форму, поскольку за счет ее из АДФ и неорганического фосфора синтезируется АТФ и запасается значительное количество энергии – 8–10 ккал/моль. Дальнейшие исследования показали, что синтез АТФ сопряжен с транспортом электронов – циклическим или нециклическим.
23. Сравнительный анализ биохимических превращений С3- и С4-путей фотосинтеза.
Цикл Кальвина (С3). Этот цикл начинается с присоединения СО2 к акцептору рибулозо-1, 5-дифосфату (РДФ). Присоединение СО2 к тому или ионному веществу называется карбоксилированием, а фермент катализирующий такую реакцию – карбоксилазой. В данной реакции карбоксилирование происходит с участием фермента рибулозодифосфаткарбоксилазы (РДФ-карбоксилаза). в присутствии воды сразу распадается на две молекулы 3-фосфоглицириновой кислоты (3-ФГК). Для дальнейших превращений ФГК необходимы АТФ и НАДФН. Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1, 3-дифосфоглицириновая кислота. Реакция катализируется ферментом фосфоглицераткиназой: Затем происходит восстановление за счет НАДФН и образуется фосфоглицириновый альдегид ФГА: Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО). Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида. Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями. ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА): Это первый путь превращения ФГА. Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА). Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК.
Это второй путь превращения ФГА. Фруктозо-1, 6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в среде неорганического фосфата. Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов. Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (транскетолаза). Затем эритрозо-4-фосфат реагирует с равным количеством ФДА и образуется седагептулозо-1, 7-дифосфат (альдолаза), которая фосфорилируется до седагептулозо-7-фосфата с участием седагептулозодифосфатазы. Четвертый путь превращения ФГА связан с его реакцией с седагептулозо-7-фосфатом с образованием равных (эквимолярных) количеств рибозо-5-фосфата и ксилулозо-5-фосфата. Ксилулозо-5-фосфат эпимиризуется, а рибозо-5-фосфат изомерезуется до рибулозо-5-фосфата, последняя фосфорилируется за счет АТФ и образуется рибулозо-1, 5-дифосфат – первичное соединение цикла Кальвина (акцептор СО2). В этих реакциях тратится еще три молекулы АТФ. Из приведенных реакций цикла Кальвина видно, что фотосинтез, являясь процессом запасания энергии, тем не менее, для своего существования требует затраты энергии. В цикле Кальвина образование фруктозо-6-фосфата можно представить в виде следующего суммарного выражения: 18 молекул АТФ запасают около 140 ккал и 12 НАДФН. АТФ и НАДФН, которые образуются в световой стадии и используются на восстановление СО2, получили название ассимиляционной силы. Цикл Кальвина подразделяют на три фазы: – карбоксилирующую РДФ + СО2 → 2ФГК; – восстановительную ФГК → ФГА; – регинирующую ФГА → РДФ.
ФЕП образуется из пировиноградной или 3-фосфоглицериновой кислоты. В результате β -карбоксилирования ФЕП превращается в четырехуглеродную щавелевоуксусную кислоту: ФЕП + СО2 + Н2О → ЩУК + Фн Фермент, который катализирует присоединение СО2 к ФЕП – фосфоенолпируваткарбоксилаза – найден сейчас у многих одно- и двудольных растений. Образовавшаяся ЩУК при участии НАДФН (продукта световой реакции фотосинтеза) восстанавливается до яблочной кислоты (малата): ЩУК + НАДФН + Н+ → малат + НАДФ+ Реакция катализируется НАДФ+-зависимой малатдегидрогеназой, локализованной в хлоропластах клеток мезофилла. Эволюционно С3-цикл появился раньше, чем С4, он есть у водорослей. В древесных растениях цикла Хетча – Слека нет. Это тоже подтверждает, что этот цикл возник позднее. В заключение отметим, что, циклы Кальвина и Хетча – Слека действуют не изолированно, а строго координировано. Взаимосвязь этих двух циклов получила название «кооперативного» фотосинтеза.
|
Последнее изменение этой страницы: 2017-04-13; Просмотров: 1017; Нарушение авторского права страницы