|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
РЕАКЦИЯ МЫШЦ НА ПАССИВНОЕ РАСТЯЖЕНИЕ
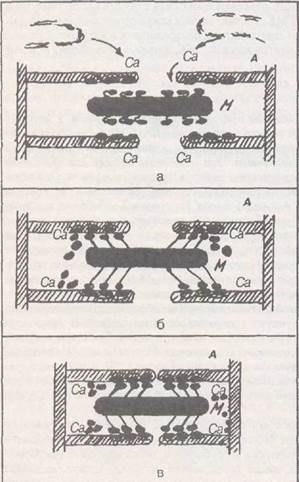
Если скелетную мышцу растягивать, то в ней возрастает напряжение, как в любой эластической структуре. Чем больше растяжение, тем выше это напряжение. У гладких мышц ситуация иная — в ответ на растяжение первоначально напряжение в мышце действительно возрастает, но затем (достаточно быстро, например, через 30—60 с) напряжение спонтанно снижается почти до исходного уровня. Таким образом, гладкая мышца ведет себя как пластическая или вязко-эластическая структура. Это свойство названо пластичностью. Если бы его не было, то мышечная стенка полых органов не смогла бы выполнять функцию резервуара. Например, при накоплении мочи в мочевом пузыре давление в нем, несмотря на то, что вместимость мочевого пузыря ограничена, не возрастает, т. к. гладкие мышцы стенки пузыря при таком растяжении постепенно снижают свой базальный тонус. СИЛА МЫШЦ Удельная сила мышц, скелетных и гладких (в расчете на 1 см2 площади поперечного сечения), почти одинакова и, в среднем, составляет Л—3 кгс или 40—30 Шсм2. Сила скелетной мышцы зависит от многих факторов. Например, от числа двигательных единиц (ДЕ), возбуждаемых в данный момент времени. Так, если мышца представлена 10 ДЕ, а в данный момент активна 1 ДЕ, то мышца способна развить силу, равную 1/10 от ее максимальной силы. Если 5 ДЕ активны, то соответственно, мышца развивает 50% от максимума и т. д., а 100% силы она разовьет в том случае, если все 10 ДЕ одновременно будут возбуждены. Сила зависит от синхронности работы ДЕ. Так, если все 10 ДЕ начнут одновременно возбуждаться, то сила будет, например, 4 кгс/см2, а если они возбуждаются асинхронно, то максимальная сила составит 3 кгс/см2. Сила мышц зависит от той частоты, с которой бегут потенциалы действия (ПД) по данным аксонам к соответствующим мышечным волокнам. Например, если альфа-мотонейрон генерирует за! секунду 20 ПД, то сила мышц будет равна 2 кгс/см2, а если 50 имп/с — 4 кгс/см2 (согласно явлению оптимума частоты раздражения, о чем говорилось выше). Сила мышцы (напряжение, развиваемое в момент ее сокращения) зависит от исходной длины. Существует некоторая средняя величина JIq (это длина мышцы при покое в условиях целостного организма), при которой мышца развивает максимальное сокращение. Если длина будет меньше Л^ или, наоборот, больше Ло (перерастянута), то сила, развиваемая мышцей в момент ее возбуждения, будет значительно меньше. Оказалось, что максимальная сила развивается мышцей в том случае, когда длина саркомера составляет 2, 2—2, 5 мкм. Зависимость силы мышцы от ее длины очень важна — особенно для сердечной мышцы (закон Франка-Старлинга) в практическом и теоретическом отношениях (она доказывает гипотезу скольжения протофибрилл, объясняющую механизм сокращения). Сила гладких мышц тоже зависит от исходной длины: существует оптимальная длина мышцы, при которой мышца развивает максимальную для нее силу. Это важный механизм саморегуляции активности гладкой мышцы. Максимальная сила гладких мышц тоже зависит от синхронности возбуждения всех ГМК, составляющих сократительный аппарат данной мышцы, от числа ГМК, вовлекаемых при действии раздражителя в акт сокращения, а также от величины входа ионов кальция внутрь каждой ГМК, который происходит при действии на ГМК веществ-стимуляторов. МЕХАНИЗМ СОКРАЩЕНИЯ Сократительный аппарат скелетной мышяы представлен миофибриллами, в которых упакованы протофибриллы — толстые и тонкие нити (филаменты). На всем протяжении миофибриллы разделены на отдельные отсеки (саркомеры), длина которых составляет в среднем 2, 5 мкм. Они ограничены Z-мембранами. Эти мембраны служат для крепления актяновых нитей. В центре саркомера расположены толстые (миозиновые) нити. Они образуют А-диск (анизотропный). Для скрепления толстых нитей имеется мембрана М, которая расположена в центре саркомера. Длина А-диска 1, 6 мкм. На уровне Z-мембран к каждому саркомеру спускается поперечная трубочка (Т-трубочка), совокупность которых названа Т-системой. Эти трубочки подходят близко (но не вплотную, щель — 10—20 им) к терминальным цистернам саркоплазматического ретикулюма. СаркоплазматическиЙ ретикулюм (СР) представлен терминальными цистернами (около Z-мембран) и продольными трубочками. СР содержит ионы кальция примерно в концентрации 10 - мМ, чего в принципе достаточно лишь для 5—6 сокращений. В момент генерации потенциала действия (ПД) происходит распространение ПД вдоль продольной плазматической мембраны, деполяризация передается на Т-трубочку, которая контактирует с терминальной цистерной. В результате открываются кальциевые каналы, по которым кальций входит в межфибриллярное пространство миофибриллы, что приводит к инициации сокращения. Потом мембраны продольных трубочек СР начинают активно откачивать ионы кальция назад в СР, и его концентрация в межфибриллярном Пространстве уменьшается, что приводит к расслаблению. В целом, описанное явление получило название электромеханическое сопряжение (ЭМС), или электромеханический каплинг. Существуют экспериментальные способы разобщения ЭМС, например, портняжную мышцу выдерживают 3—4 часа в растворе 400 ммоль/л глицерина, а затем ее помещают в обычный раствор Рингера. В ней «отрываются» Т-трубочки, и такая мышца «прекрасно» генерирует ПД в ответ на раздражение, но не сокращается. Кофеин, кстати, вызывает высвобождение кальция, минуя эту систему сопряжения. В настоящее время в практике используется миорелаксант типа дантрилена, который разобщает обратимо ЭМС и, тем самым, вызывает миорелаксирующий эффект. Было показано, что утомление изолированной мышцы может наступать в результате разобщения ЭМС. Недавно было отмечено, что утомление вызывается тем, что кальций перестает выходить из терминальных цистерн СР. Относительно механизма, с помощью которого происходят передача деполяризации с Т-трубочки на терминальные цистерны (своеобразный внутриклеамшый синапс), сущест-
вуют разные точки зрения. Во-первых, в щелях между Т-трубочкой и терминальными цистернами обнаружены мостики-каналы (шириной до 15 нм), которые, возможно, представляют собой «затычку» для кальциевых каналов. Полагают, что деполяризация убирает «затычку» и тем самым повышает проницаемость кальциевых каналов терминальной цистерны. Это так называемая механическая гипотеза. Согласно электрической гипотезе, деполяризация сама по себе настолько сильна в Т-трубочках, что этого достаточно для открытия потенциал зависимых кальциевых каналов терминальной цистерны. По химической гипотезе, между деполяризацией в Т-трубочках и открытием кальциевых каналов терминальной цистерны имеется посредник — это ионы кальция или инозитол-трифосфат. Кстати, недавно обнаружено, что в Т-трубочках действительно есть свои собственные кальциевые каналы, которые могут при определенных условиях породить кальциевые ПД. Возможно, эти самые каналы впускают ионы кальция к терминальной цистерне, а эти ионы как затравка открывают собственные кальциевые каналы мембраны терминальной цистерны.
Вошедшие в межфибриллярное пространство ионы кальция инициируют сокращение. Это происходит в результате того, что кальций соединяется с одной из 3 субъединиц (кальций-связующая субъединица) молекулы тропонина, которая располагается с интервалом в 40 нм на ак-тиновой нити. В условиях покоя эта молекула тропонина создает условия для того, чтобы фибриллярная молекула тропомиозина, находящаяся на актиновой нити, не давала миозино-вому мостику контактировать с актином (в условиях покоя мостик не может соединиться с актиновой нитью, т. к. ему мешает тропомиозин). Когда же кальций соединится с тро-понином, то происходят конформа-ционные изменения в другой субъединице тропонина (ингибирующая субъединица), в результате чего нить тропомиозина продвигается в глубину бороздки и освобождает места на актиновой нити для связывания с миозином. Описанная система получила название актин-связующая регуляция. Тропонин и тропомиозин называются регулирующими белками, т. к. они регулируют состояние в саркомере актиновых и миозиновых нитей. Сокращение происходит, согласно распространенной модели А. Хаксли (модель скольжения или, как сейчас называют — «модель скользящий филамент — вращающийся мостик» — А. Хаксли, Симмонс, 1971), за счет скольжения актнновых нитей в промежутках между миозиновыми. Этот процесс осуществляется поперечными мостиками миозина. Поперечные мостики представляют собой головы миозина (миозин состоит из 2 субъединиц — легкий меромиозин — хвост миозиновой нити и тяжелый меромиозин). Тяжелый меромиозин заканчивается субъединицей С-1 — головкой миозиновой молекулы. Головка соединена с телом миозиновой нити с помощью субфрагмента С-2 (шейка), который может изгибаться. Головка обладает АТФ-азной активностью, которая проявляется, однако, в присутствии актина, когда головка будет непосредственно прикреплена к актиновой нити. В условиях покоя мостик (головка) не прикреплен к актиновой нити — мешает тропо-миозин. На кончике мостика находится молекула АТФ. Когда появляется кальций и отодвигается тропомиозин от актиновой. нити (снимается экранировка), мостик под углом 90° цепляется к актиновой нити. Тут же происходит активация АТФ-азной активности и как следствие — гидролиз АТФ с выделением порции энергии. Эта энергия используется для того, чтобы создать крутящий момент («гребок»), в результате которого мостик проталкивает актиновую нить примерно на 10 нм (это меньше 1% длины саркомера). Если рядом с мостиком имеется свободная молекула АТФ, то она встраивается на вершину мостика и обеспечивает отрыв мостика от актиновой нити. Если в среде много кальция, то актиновая молекула по-прежнему свободна от экрана (от тропомиозина), и потому мостик вновь прикрепляется к нити, но уже в другом месте, и вновь повторяется цикл. За период укорочения (напряжения) мостик успевает совершить 50 циклов («гребков»), в результате чего длина саркомера уменьшается примерно на 50%. Если уровень кальция снижается (в результате активности кальциевого насоса и прекращения выхода кальция из терминальной цистерны), то наступает процесс расслабления (удлинения). В случае, когда АТФ исчерпана, наступает ритор мышцы — нет расцепления между актиновой и миозиновой нитями. Это имеет место, например, при трупном окоченении. В скелетных мышцах запас АТФ невелик — всего на 10 одиночных сокращений. Поэтому необходим постоянный ресинтез АТФ. Существуют три пути. 1) Ресинтез АТФ за счет креатинфосфата (КФ), запасы которого ограничены. Реакция идет очень быстро, поэтому можно за несколько секунд совершить огромную работу, что и делается, например, спринтером или штангистом, когда он совершает рывок. Но ограниченность запасов приводит к тому, что ресинтез АТФ в анаэробных условиях долго идти не может. 2) Гликолитичес-кий путь ресинтеза: он связан.с анаэробным расщеплением глюкозы до молочной кислоты. В результате образуется 2 моля АТФ на 1 моль глюкозы. Этот путь тоже достаточно мощный, идет быстро, но из-за того, что молочная кислота, которая при этом накапливается, тормозит активность гликолитических ферментов, его возможности тоже ограничены. Обычно этот вид ресинтеза АТФ совершается в пределах 20—120 секунд. Поэтому он используется при беге на средние дистанции (например, 200, 400, 800 м). Замечено, что этот вид ресинтеза всегда имеет место в начале всякой двигательной активности, пока кровообращение в работающей мышце не станет адекватным для проведения 3-го типа ресинтеза АТФ. 3) Это аэробное окисление глюкозы и жирных кислот в цикле Кребса. Он совершается в митохондриях. В среднем на 1 моль глюкозы образуется около 38 моль АТФ, при окислении 1 моля жирной кислоты — около 128 моль АТФ. Этот процесс очень экономный, однако для получения таким образом энергии требуется больше времени, чем при первых двух способах. Поэтому 3-й путь ресинтеза используется во всех случаях, где мощность работы невысокая. В обыденной жизни именно этот путь наиболее широко эксплуатируется нашими мышцами. А запасы углеводов (гликоген, свободная глюкоза) и жиров (источник жирных кислот) достаточно велики. Например, за счет окисления только гликогена человек может непрерывно пробежать 15 км пути; запасов жиров так много, что их хватит на несколько недель непрерывной работы. |
Последнее изменение этой страницы: 2017-05-04; Просмотров: 669; Нарушение авторского права страницы