
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
РЕФЛЕКТОРНЫЙ ПРИНЦИП РЕГУЛЯЦИИ
Идею о том, что организм, наделенный нервной системой, способен отвечать на действие внешних раздражителей по типу «кнопка — ответ», высказал французский философ Рене Декарт (XVII в.). Термин «рефлекс» был введен Иржи Прохазкой (конец XVIII в.). Многие выдающиеся физиологи XIX и XX вв. разрабатывали теорию рефлекторной деятельности. Огромный вклад внесли И. М. Сеченов, И. П. Павлов. По мнению И. М. Сеченова, все сознательное и бессознательное совершается по типу рефлекса (рефлекс-отражение). Ему также принадлежит важная идея о том, что рефлекторная деятельность осуществляется с участием тормозных процессов в ЦНС (он открыл само явление торможения в ЦНС). И. П. Павлов, разделяя идею о рефлексе как основе деятельности ЦНС, выделил особый класс рефлексов, который он назвал «условные рефлексы», подробно их изучил и сформулировал важное положение о том, что в основе высшей нервной деятельности человека и животных лежат условные рефлексы, которые формируются на базе безусловных рефлексов за счет появления временных связей. П. К. Анохин считал, что все многообразие деятельности человека и животного можно объяснить с помощью концепции функциональных систем (ФС)> механизмы которых лежат в основе целенаправленной деятельности. Атрибутами функциональных систем служат: афферентный синтез, принятие решения, эфферентный синтез, акцептор результата действия. В каше время идет поиск «новых» (точнее — иных) форм деятельности ЦНС, позволяющих объяснить принципы работы мозга. С этих позиций предлагается, например, представление о существовании комплекса фиксированных действий: это объединение нейронов (нервный центр)» в которых отсутствует афферентный нейрон (или может отсутствовать), но обязательно есть пейсмекерный и эф- ферентный нейроны. Независимо от наличия во внешней среде стимула этот центр работает в автоматическом режиме: пейсмекер задает последовательность возбуждений эфферентному нейрону, а тот посылает в соответствии с заданным тактом сигналы-команды к эффектору. Очевидно, такая ситуация имеет место в дыхательном центре и в сшшальных механизмах, обеспечивающих шаговые движения. Полагают, что комплексы фиксированных действий контролируются другими отделами ЦНС. Шеперд Г. считает, что имеются и так называемые «центральные системы», которые осуществляют сложные регуляторные процессы ради получения определенной цели. Эта идея похожа на концепцию П. К. Анохина о функциональных системах. Итак, вряд ли ЦНС «справляется» со своими управленческими задачами только с помощью рефлекса. Но пока в курсе нормальной физиологии работает лишь идея «рефлекса».
Самая простая рефлекторная дуга — моносинаптическая. Она состоит из 2 нейронов: афферентного и эфферентного. Обычно латентный период, т. е. время от момента нанесения раздражителя до конечного эффекта (или это называется временем рефлекса) — достигает в таком случае 50—100 мс, а центральное время — промежуток времени, в течение которого импульс пробегает по структурам мозга, составляет около 3 мс. Известно, что для прохождения 1си-напса в среднем требуется око- ло 1, 5 мс. Таким образом, центральное время рефлекса косвенно указывает на число синап-тических передач, имеющих место в данном рефлексе. Моносинаптические рефлексы — это сравнительно ограниченный круг рефлексов. Классический пример — спинальные мио-татические (возникающие в ответ на растяжение мышцы) рефлексы. Чаще дуга рефлекса представлена 3 и более последовательно соединенными нейронами — афферентным, вставочными и эфферентным. Центральное время у таких рефлексов больше 3 мс (например, если 2 синаптических переключения — то около 4—6 мс). В целом лолисинаптические рефлексы — достаточно широко распространенные виды рефлексов. Например, сгибатель-ный рефлекс, возникающий при раздражении рецепторов кожи. Виды рефлексов. Отметим наиболее существенные моменты этой классификации. 1. Безусловные и условные рефлексы — по способу образования рефлекторной дуги. 2. Моносинаптические, лолисинаптические — по компонентам рефлекторной дуги. 3. Спинальные, бульбарные, мезенцефальные, кортикальные — по расположению ос 4. Интерорецептивные, экстерорецептивные — по характеру рецепторов, раздражение 5. Половые, оборонительные, пищевые и т. д. — по биологическому значению рефлекса. 6. Рефлексы соматической и вегетативной нервной системы (или — соматические, веге 7. Сердечные, сосудистые, слюноотделительные — по конечному результату. ТОРМОЖЕНИЕ В ЦНС. ЕГО ВИДЫ И ПРЕДПОЛАГАЕМЫЕ МЕХАНИЗМЫ
Впервые идею о том, что в ЦНС помимо процессов возбуждения существует процесс торможения, высказал И.М. Сеченов, Исследуя рефлекторную деятельность ля-гушки с сохраненными зрительными буграми, И.М. Сеченов определял время сгибательно-го рефлекса (по методике Тюрка) — в ответ на погружение лапы в кислоту происходило сгибание конечности в тазобедренном и коленном суставах. Если на зрительный бугор поместить кристаллы соли, то возникает торможение — удлинение времени рефлекса. Это наблюдение и позволило И.М. Сеченову говорить о явлении торможения. В после- дующем такой вид торможения получил название сеченовское торможение, или центральное торможение. . Гольц наблюдал, что торможение можно получить^ прикладывая дополнительный раз В ЦНС огромное число тормозных нейронов. Некоторые из этих нейронов имеют «собственное» имя — в честь открывателя этих структур, например, клетки Реншоу, клетки Уилкинсона и т. д. По-видимому, каждый тормозной нейрон вырабатывает какой-то один тип медиатора (например, глицин или гамма-аминомасляную кислоту, FAMK) и осуществляет в соответствии с этим торможение. Различают два вида торможения — пресинаптиче-ское и постсинаптическое. Пресинаптическое торможение осуществляется путем выторма-живания какого-то определенного пути, идущего к данному нейрону. Например, к нейрону подходят 10 аксонов, и к каждому из этих аксонов подходят аксоны от тормозных нейронов. Они могут тормозить проведение соответственно по каждому из аксонов в отдельности. Процесс лресинаптического торможения протекает по типу католической депрессии: в области контакта выделяется ГАМК, которая вызывает стойкую деполяризацию, что нарушает проведение волны возбуждения через этот участок. Второй тип— постсинаптическое торможение (за счет гиперполяризующего действия глицина) — происходит в результате гиперполяризадии всего нейрона, поэтому блокируется весь нейрон одновременно. Б локатором ГАМК-ергических рецептов является бикукулин, а блокатором глициновых рецепторов — стрихнин, столбнячный токсин. Если рассмотреть «архитектуру» использования тормозных нейронов при организации нейронных сетей, цепей и рефлекторных дуг, то можно выделить ряд вариантов этой организации (это отражается в названии данного вида торможения)... 1. Реципрокное торможение. Как пример: сигнал от мышечного веретена поступает с 2. Возвратное торможение. Альфа-мотонейрон, к примеру, посылает аксон к соответст 3. Существует ряд вариантов возвратного торможения, в частности, латеральное тормо |
Последнее изменение этой страницы: 2017-05-04; Просмотров: 522; Нарушение авторского права страницы

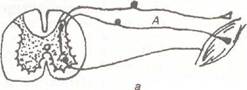 Рефлекторная дуга — это морфологическая основа рефлекса. Рефлекс — это закономерная реакция организма на действие раздражителя (обратите внимание — на действие раздражителя, а если его нет — значит, это не рефлекс) при обязательном участии ЦНС. Различаются также рефлексы, реализующиеся через клетки нервных ганглиев. В любом случае должна быть рефлекторная дуга. В случае безусловных рефлексов она формируется независимо от жизненного опыта индивидуума — становление безусловных рефлекторных дуг генетически запрограммировано. Ус-
Рефлекторная дуга — это морфологическая основа рефлекса. Рефлекс — это закономерная реакция организма на действие раздражителя (обратите внимание — на действие раздражителя, а если его нет — значит, это не рефлекс) при обязательном участии ЦНС. Различаются также рефлексы, реализующиеся через клетки нервных ганглиев. В любом случае должна быть рефлекторная дуга. В случае безусловных рефлексов она формируется независимо от жизненного опыта индивидуума — становление безусловных рефлекторных дуг генетически запрограммировано. Ус- ловно-рефлекторный процесс требует создания новых рефлекторных дуг на основе сочетания работы безусловных рефлекторных дуг и индифферентного раздражителя.
ловно-рефлекторный процесс требует создания новых рефлекторных дуг на основе сочетания работы безусловных рефлекторных дуг и индифферентного раздражителя.