|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
IX этап. Вылупление (220 сут).Стр 1 из 6Следующая ⇒
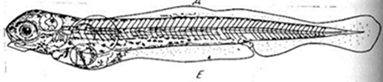
IX этап. Вылупление (220 сут). Зародыши освобождаются из оболочек не все одновременно. Выклев начинается с появления единичных личинок, затем количество их увеличивается. Момент, когда в аппарате уже имеется несколько десятков личинок, можно считать началом выклева. При хороших условиях развития число личинок быстро увеличивается, и наступает массовый выклев. Освобождение зародышей от оболочек происходит следующим образом. В железе вылупления зародыша образуется особое вещество, которое с приближением стадии выклева начинает выделяться из железы вылупления в перивителлиновое пространство. Выделение фермента происходит постепенно и приводит к перевариванию оболочек, в результате снижается их прочность. Двигаясь в ослабленных оболочках, зародыш растягивает, а затем и разрывает их. Сначала появляется голова личинки или хвост, а потом освобождается вся личинка. Свободный эмбрион. Запасы желтка сильно сокращены. Печень позади желточного мешка. Грудные плавники сильно увеличены
Таблица 6. Эмбриональный период развития омуля
Комбинированный метод Комбинированный способ выращивания сеголетков сигов разделяется на 2 этапа. На первом этапе продолжительностью около месяца молодь выращивается в бассейнах. На втором этапе ее выращивание до сеголетка осуществляется в садках. Преимущество этого метода заключается в следующем: - при продолжении выращивания молоди в бассейнах исключается лишнее ее перемещение, а следовательно, и травмирование; - сохраняется возможность регулирования условий выращивания (температуры, освещения, газового режима, водообмена и др.); - обеспечивается тщательный контроль за состоянием выращиваемой молоди; - при выращивании молоди в садках исключается ее перемещение в новые условия для получения товарной продукции; - снимается стрессовая ситуация сеголетков, возникающая при их перемещениях; - обеспечивается подготовка молоди к условиям производства товарной продукции; - не снижается темп роста при переводе сеголетков на новый этап развития; - сохраняется высокая выживаемость молоди на всех этапах развития (малек, сеголеток, годовик и более старшие возрастные группы). К началу малькового периода личинки сига обычно достигают средней массы около 40 мг при средней длине около 19 мм. Дальнейшее выращивание молоди до массы тела 200 мг целесообразно продолжить в тех же рыбоводных емкостях, используя как круглые (диаметр — 1,0-1,2 м, глубина — 0,5-0,6 м), так и квадратные (1,5х1,5х0,5 м) бассейны. Начальная плотность посадки молоди — 30 тыс. особей/м3, конечная плотность — около 8,0 тыс. особей/м3, то есть в процессе роста молоди плотность ее посадки необходимо уменьшать. Благоприятная температура воды — 10-16 °С. Проточность воды в бассейнах около 4 л/мин. Содержание кислорода на вытоке не менее 7 мг/л. В это время используется гранулированный корм ГосНИОРХа — МС-84, ВНИИПРХа — РГМ-СС, Raisio agro — Nutra ХР 0,5, Royal 1,2 и др. Размер гранул не должен превышать 0,3 мм. При больших размерах гранул корм для молоди этой массы тела будет недоступен. Частота кормления должна быть 6-8 раз в сутки. Конечно, частоту кормления можно сокращать, например, до 4 раз в сутки, но тогда его поедаемость будет снижаться. Следствием этого станет сокращение усвояемости корма и замедление темпа роста. Конечно, ухудшится качество выращиваемой молоди. Норма внесения корма при увеличении температуры воды от 10 до 16 °С соответственно будет возрастать от 5,5 до 7,5 % от биомассы выращиваемой молоди. Дальнейшее выращивание молоди до средней массы 2 г можно проводить в тех же рыбоводных сооружениях или в бассейнах, размеры которых увеличиваются в 2 раза. Плотность посадки — от 8 тыс. до 1,5 тыс. экз/м3. Благоприятная температура воды — 12-17 °С. Проточность — 5 л/мин. Содержание кислорода — не менее 7 мг/л (на вытоке). Корма МС-84, РГМ-СС, Nutra ХР 0,7, Royal 1,2 и др. Размер гранул — до 0,6 мм. Использование мелких гранул потребует большей затраты энергии на их поедание и, соответственно, сократит эффективность использования пищи на рост молоди. Более крупные гранулы корма будут недоступны молоди этого размера. Режим кормления — 4-6 раз в сутки. При выращивании молоди сигов от 2 до 3 граммов можно использовать такие же рыбоводные емкости (d — 1,2 м, глубина — 0,6 м, и 1,5×1,5×0,5 м). Плотность посадки — 1,0-1,5 тыс. экз./м3. Благоприятная температура воды — 12-18 °С. Особое внимание следует обращать на водообмен в бассейнах, изменением объемов которого можно регулировать не только температуру воды, но и газовый режим. Расчетная величина проточности — около 6 л/мин. При этой величине и содержании кислорода на втоке около 8 мг/л на вытоке его показатель не снизится менее 7 мг/л, что является нижним пределом для нормального развития молоди сигов. Корма ПС-95, РГМ-СП, Nutra ХР 0,7, Royal 1/2/1/5 и др. Рекомендуемый размер гранул — до 0,8 мм. Режим кормления — 4-6 раз в сутки. При благоприятных условиях выращивания молодь сигов достигает массы тела 3 г в течение 45-55 суток. Выживаемость не менее 75 %. Молодь средней массой 3 г для дальнейшего выращивания пересаживают в делевые садки. К этому времени молодь хорошо адаптирована к изменяющимся условиям окружающей среды, чешуйчатый покров полностью покрывает все тело, хорошо развиты зрительные рецепторы. Она очень активна, интенсивно питается и быстро накапливает массу тела. В основном преобладает весовой рост. Выращивать молодь сигов массой более 3 граммов целесообразно в садках размером 3х4х4 м или 4х4х4 м. В таких садках с ячеей дели 5-6 мм более интенсивно происходит водообмен, что сохраняет нормальный газовый режим, устойчивую температуру и более эффективно выводит конечные продукты метаболизма рыб. Сохранению благоприятного режима в садках также способствует наличие проточности (не более 0,05 м/с). При этом сохраняется возможность более тщательного наблюдения за поведением и состоянием выращиваемой молоди. Плотность посадки 800-1000 экз./м3. При достижении молодью массы 10 г плотность посадки следует сократить до 800-400 экз./м3, а более 25 г — до 300-250 экз./м3. При этом размеры ячеи в дели увеличиваются до 6-10 мм. Благоприятная температура воды — 12-22°С, оптимальная — 14-18°С. Содержание кислорода больше 7 мг/л. Корма РГМ-СП, ПС-95, Nutra starter-1, Nutra parr-1.2 и др. Режим кормления 3-4 раза в сутки. По мере роста молоди возможно кормление 2-3 раза в сутки. При 2-разовом кормлении следует тщательно следить за поедаемостью корма и ростом молоди. Раздача и дозирование корма более эффективны при использовании автоматических кормушек. В настоящее время существует множество таких кормушек. Поэтому рыбовод должен подобрать такую конструкцию, использование которой наиболее эффективно в его хозяйстве. На этапе садкового выращивания сеголетков необходимо следить за чистотой делевых стенок. Регулярная (раз в 2-4 суток) чистка садков будет обеспечивать благоприятный режим для выращиваемой молоди. Профилактические мероприятия (не реже одного раза в 1,5 месяца) позволят сохранить здоровье молоди и получить качественный посадочный материал. Все оборудование для ухода за рыбой и садками должно быть индивидуальным. Соблюдая технологию выращивания и сохраняя благоприятные условия, за три месяца можно получить сеголетков массой 30-35 г. Максимальная выживаемость составляет 80 %. Выращивание рыбы в озерных хозяйствах Сбор и инкубация икры Преднерестовую сортировку производителей проводят в садках при температуре воды 3,5-4°С. Особей, не созревших в этот сезон, перемещают на зимовку в выростные садки. Созревающих самок и самцов размещают раздельно в садки диаметром 4-5 м с глубиной погружения 2,5-4 м. Просмотр на зрелость чира и муксуна начинают при температуре воды до 2,5°С через два дня, а пеляди - через три дня. Первую порцию спермы не берут, так как она обычно бывает низкого качества. Оплодотворение производят полусухим способом. Соотношение самок и самцов составляет 1 : 2. Самцов используют многократно с промежутком 4-6 дней. При снижении температуры воды в садках до 3 °С проводят осмотр самок. Зрелых самок переносят в рыбоводное помещение, где от них отцеживают икру. Массовое созревание самок наблюдается в первой половине декабря при температуре воды 0,2-0,3°С. Только баунтовские сиги мечут икру подо льдом в конце марта - начале апреля. Пелядь речная созревает перед ледоставом в октябре при температуре 2°С и ниже, озерная форма - в конце ноября-декабре при температуре 0,2-0,8°С. Европейская и сибирская ряпушки, муксун, чир, сиг-лудога, чудской, пыжьян нерестуют в момент ледостава. При этом чир выметывает икру только в проточной воде. Икру получают в помещении. После получения икры самок выдерживают 2-3 дня в отдельном садке для выявления сильно ослабевших и травмированных, после чего их объединяют и выращивают до следующей нерестовой кампании. Инкубация продолжается всю зиму в течение 6 месяцев. Икру и сперму берут только от производителей с текучими половыми продуктами, которые выделяются уже при слабом нажатии на брюшко. Икру, среди которой встречаются икринки светло-серого и белого цвета, не берут. В один таз отцеживают икру от 2-3 самок и добавляют сперму от 1-4 самцов, затем икру тщательно перемешивают и оставляют в покое на 4-5 мин, затем добавляют еще воды и вновь перемешивают. После этого приступают к промывке и обесклеиванию икры в большом объеме воды, сменяя воду 20-30 раз. Процесс набухания у сиговых протекает в спокойном состоянии на протяжении 1,5-6 ч. Конечная прочность оболочки икры наступает через 7-14 ч, слой воды в тазу с набухающей икрой составляет 15-20 см. Воду меняют через 30-40 минут. Объем икры в период набухания увеличивается на 150%. Оплодотворенную и освобожденную от клейкости икру после набухания при необходимости перевозки загружают на рамки, которые стопкой устанавливают в специальных транспортировочных ящиках. Длительность транспортировки в первые два дня после оплодотворения не должна превышать 12 ч. На стадии морулы икру можно перевозить в течение 2-3 суток, т. е. на 3-5-е сутки при температуре 2-5°С. Икра в этот период более устойчива к колебаниям температурного и кислородного режимов. Перевозка икры должна быть закончена в конце стадии дробления, которая наступает через 7-10 суток (около 500-700 градусодней). Доставленную икру после адаптации к температуре и учета загружают в аппараты Вейса (тыс. шт): для ряпушки, рипуса - до 900, пеляди - 800, сигов (чудского, лудоги, пыжьяна) - до 300, омуля байкальского - до 300, чира, муксуна, нельмы - до 200. В процессе инкубации расход воды в аппаратах регулируют. В начале инкубации (три дня) и в конце вылупления расход составляет 0,05 л/с (3 л/мин). В период отбора расход воды уменьшают до 0,04 л/с (2,2 л/мин). Температура воды в период инкубации поддерживается на уровне 0,2-0,8°С. Отбор мертвой икры проводят через каждые 10-20 суток развития. Отбору подлежит поврежденная икра, неоплодотворенная и аномальная в развитии белого цвета. Неоплодотворенная икра составляет 8-16%. Общий отход икры за период инкубации может достигать 50%. Затем отход икры наблюдается на стадии гаструляции и особенно на стадии замыкания бластопора. В это время не допускают перемешивания икры в аппарате. Икру из аппарата отсасывают сифоном. Большой урон инкубируемой икре наносит развитие гриба - сапролегнии, от которого необходимо своевременно освобождаться. Для дезинфекции икры в случае появления очагов сапролегнии ее промывают в контрольных аппаратах раствором малахитового зеленого в концентрации 1 : 180000 или формалином в концентрации 1 : 2000. Для инкубации икры сиговых используют чистую низко- или среднеминерализованную воду карбонатного класса без взвесей ила, песка, не загрязненную сточными водами и нефтепродуктами (табл.). Полный химический анализ воды в период инкубации проводят 3-4 раза. Таблица. Химический состав воды, необходимый в инкубационно-личиночном сиговом цехе
Отход икры сиговых дополнительно к количеству неоплодотво- ренной и погибшей при сборе и транспортировке икры составляет в норме (в %): для чира - 25, озерной пеляди - 15, речной пеляди - 12, сибирской ряпушки - 8. Неблагоприятное воздействие факторов среды на последних стадиях развития может вызвать раннее вылупление. Массовое вылупление эмбрионов чира наблюдается при весеннем повышении температуры воды до 3-4°С, муксуна - до 4-6°С, пеляди — до 5-8°С. При нормальных условиях инкубации икры, полученной от производителей хорошего качества, количество дефектных эмбрионов не превышает 4,0%. Рыбоводно-технологические нормы инкубации икры представлены в таблице. Таблица. Биотехнологические нормы инкубации икры сиговых
После вылупления эмбрионы поднимаются в верхние слои аппарата Вейса и по желобу с током воды выносятся в уловитель, где концентрируются. По мере накопления личинок отчерпывают ведром и переносят в непроточный отстойник, где оболочки икры оседают на дно и их оттуда отбирают сифоном. Затем личинок помещают в лотки для выдерживания и подращивания. Выдерживание личинок проводят также в сетчатых садках из газа № 13-17 при постоянной проточности и температуре воды 1-2°С для чира и муксуна, 4-8°С - для пеляди и ряпушки. Плотность выдерживания свободных эмбрионов и проточность зависят от температуры воды и содержания в ней растворенного кислорода. Вода в лотки и бассейны для выдерживания личинок подается снизу при верхнем сливе. Вода должна быть чистой, без механической взвеси и пузырьков воздуха. Освещение лотков должно быть обязательно равномерным. Выдерживание продолжается до перехода свободных эмбрионов на внешнее питание (до момента рассасывания желточного мешка). Обычно этот процесс длится 3-4 дня, но при пониженной температуре воды процесс удлиняется. Поэтому следует поддерживать температуру воды в пределах 8-10°С. Содержание зоопланктеров в этой воде должно быть до 300-400 экз./л. В начальный период развития движение личинок носит мерцательный характер в вертикальном направлении и ограничивается небольшим пространством. Иногда личинки образуют большие скопления. Плотность посадки при выдерживании их в лотках составляет 400-500 шт./л. При этом в лотках должна быть постоянная проточность, а полный водообмен осуществлялся не реже 2-2,5 ч. Увеличение времени водообмена до 3,5-4 ч приводит к снижению кислорода до 5-6 мг/л, что является нижним пределом. Содержание свободной углекислоты (СО2) в лотках при выдерживании личинок не должно превышать 8-10 мг/л, поэтому лотки не должны быть глубже 0,4-0,5 м. Ширина и длина их могут быть различными. Стенки должны быть гладкими, а половина дна выложена белой плиткой, чтобы хорошо просматривалась толща воды. В таких условиях личинок можно выдерживать на заводе без подкормки только до личиночной стадии развития и рассасывания желточного мешка на 50-60 %, то есть, при температуре воды 2-3 °С 3-4 дня. Эмбрионы речной и озерной пеляди выклевываются в апреле - мае, когда водоемы вскрываются ото льда. Свободных эмбрионов выдерживают в лотках цеха с проточной водой 3-5 дней, в течение которых они достигают личиночной стадии развития, после чего их высаживают в озеро. Эмбрионы чира, гибридов чир × пелядь и пелядь × чир выклевываются в конце марта - середине апреля. До вскрытия озер их выдерживают на заводе при температуре 1-2 °С. В течение этого времени они достигают стадии личинки. При повышении температуры воды до 4-6 °С и рассасывании желточного мешка наполовину, личинки переходят на активное питание живым кормом. В это время расход корма составляет 20-30 % от массы личинок. Хороший рост личинок отмечен при кормлении их смешанным кормом: молодь моин и науплиусами артемии. Выживаемость личинок - 80-85 %. При подращивании личинок в лотках до малькового периода развития их плотность посадки снижается до 100 шт./л, затем до 50-70 шт./л. Основным фактором, определяющим рост и развитие личинок, является температура воды. Оптимальная температура воды при выращивании личинок равна 10-15 °С. Вторым фактором, положительно влияющим на рост личинок, является наличие живого корма. При отсутствии живого корма временно для подкормки можно использовать желточный порошок и сухие дафнии. Выращивание товарной рыбы Выращивание товарной рыбы является конечном этапом технологического цикла озерного рыбоводства. Практикуется двухлетний нагул сиговых рыб в озерах заморного и незаморного типов, что представляет собой эффективную технологию получения товарной продукции. Товарных рыб выращивают в поликультуре, чтобы полнее использовать кормовые ресурсы озера. В состав поликультуры обычно входят рыбы-планктофаги, бентофаги, хищники и растительноядные. Поликультура позволяет эффективно использовать все зоны в водоеме (пелагиаль, профундаль, литораль). Например, в озерах Северо-западного региона основным объектом, нагуливающимся в пелагиали является пелядь. Целесообразность использования хищных рыб зависит от наличия в озере малоценных рыб. В хорошо облавливаемых озерах, где можно управлять численностью рыб промысловых размеров, как правило, наблюдается увеличение численности молоди (благодаря снижению смертности после отлова части рыбы промысловых размеров и хищников), рекомендуется включать в состав поликультуры судака. В озерах, где нет щуки, можно выращивать таких хищников, как кижуч, радужная форель и нельма. Для эффективного использования кормовой базы озера следует вселять рыб с разным характером питания и соблюдать нормы посадки. При выращивании товарных сигов в озерах к ним предъявляются определенные требования, которые показаны в таблице. Таблица. Характеристика нагульных озер для сиговых рыб
Для получения товарной рыбы масса сеголетков сиговых рыб при посадке в озера с естественным составом ихтиофауны должна быть не менее 20-25 г. Это способствует выживанию молоди под воздействием хищников, и быстрому достижению рыбой товарной массы. При выпуске на нагул в озера сеголетков хищных рыб (судака, нельмы) в первой половине лета, когда они переходят на хищный образ жизни, масса молоди может быть небольшой - 3-5 г. Щуку выпускают даже на стадии малька, равномерно распределяя вдоль береговой линии, для обеспечения ее кормом, убежищами и предупреждения каннибализма. При выращивании сигов, одним из способов борьбы с молодью местных рыб, является включение в поликультуру кубенской нельмы. Ее сеголетки к осени достигают массы 90-100 г. Половая зрелость наступает у самцов в возрасте 4 лет при длине тела 60-65 см и массе 2,0-2,5 кг, а у самок - в возрасте 5 лет при длине тела 70-75 см и массе 3,5-4,0 кг. По характеру питания кубенская нельма является типичным хищником. Молодь до двух месяцев питается зоопланктоном, затем переходит на питание личинками, а затем и молодью рыб. Пищей взрослых рыб служат мелкий окунь, ерш, молодь плотвы. Плотность посадки рыб на нагул следует рассчитывать по продукции кормовых организмов. Расчет проводят отдельно для рыб с разным характером питания по формуле: N = (KP-P)/(CP-V), где: N - коэффициент посадки рыб, шт./га; Кр - коэффициент, показывающий степень использования, запасов (долю потребления) пищи рыбами; Р - реальная продукция кормовых организмов за вегетационный период, кг/га; Ср - оптимальный рацион одной рыбы, обеспечивающий достижение запланированной массы, кг; V - коэффициент промыслового возврата от посадки. Для определения плотности посадки рыб в озере с естественным составом ихтиофауны необходимо определить пищевые потребности местных рыб, оставшихся в озере после интенсивного облова. Тогда уравнение расчета плотности посадки приобретает вид: N=Kp-(P-Ca)/Cp-V, где Са - суммарный рацион рыб-аборигенов по данному виду кормовых организмов. Расчет плотности посадки можно вести по величине потребления кормов рыбами в рыбопитомных озерах (табл.). Таблица. Суммарное потребление кормов в разнотипных озерах
Например, в мезотрофном озере местные рыбы потребляют 1000 кг/га (Са) корма за вегетационный сезон (за год), в том числе зоопланктона 400 (Са зоопланктона) и зообентоса 600 (Са зообентоса) кг/га. В подготовительный период величина изъятия малоценных рыб составила 70 %, т. е. для разводимых рыб должно было высвободиться соответственно 280 и 420 кг/га (Сав) зоопланктона и макрозообентоса. Однако в результате увеличения индивидуального рациона у 30 % оставшихся рыб (на 30-50 %) и увеличения численности их молоди в несколько раз на следующий год вселенцы смогут потребить значительно меньше корма: Кр составит не более 15-30 %, т. к. Ка достигает 85 % в неудобряемых мелководных и 70 % в удобряемых хорошо обловленных (70 % запаса) озерах. В мезотрофные озера для их эфтрофии следует вносить не более 50-60 кг/га азотно-фосфорных удобрений за вегетационный сезон. Известно, что на каждые 5-10 кг внесенных азотно-фосфорных удобрений получают 1 кг рыбы. Для питомников это соотношение 5 : 1, для нагульных озер 10 : 1. Коэффициент выживания (промыслового возврата за ряд лет) для сеголетков сиговых рыб в хорошо подготовленных озерах достигает 0,5-0,7, для двухлетков карпа-0,68-0,85. В зависимости от массы товарной рыбы меняется и величина оптимального рациона вселенцев. Например, для двухлетков сиговых массой 250-300 г суммарной рацион за сезон составляет около 2 кг, для трехлетков карпа - 2,5 кг. Таким образом, 50-60 кг/га удобрений позволяют получить 5-10 кг рыбы, потребляющей до 100 кг/га корма при кормовом коэффициенте не менее 10. Соответственно, для расчета плотности посадки на нагул рыб-зоопланктофагов (N3) таков: = (280 + 100) • (1-0,7) /2 • 0,5 = 184 шт./га. Следовательно, плотность посадки рыб-зоопланктофагов (пеляди) в удобряемые мезотрофные озера после интенсивного облова малоценных рыб составляет около 185 шт./га, при промысловом возврате 50 % будет выловлено около 100 шт./га или примерно 30 кг/га (при массе двухлетков около 300 г). Основным показателем условий нагула рыб является темп роста. О нем судят по упитанности и обеспеченности пищей, по биомассе кормовых организмов и по гидролого-гидрохимическому режиму. Товарные сеголетки пеляди должны к концу вегетационного периода достигать массы не менее 100 г, двухлетки - 250-400 г. В озерах со средней глубиной 5-10 м хороший темп роста у пеляди наблюдается с июня по август, т. е. в период наиболее интенсивного развития зоопланктона. Максимум накормленности пеляди наблюдается при температуре воды 15-16°С. Трехлетки пеляди при нормальных условиях нагула достигают массы 450-650 г, при разреженной плотности посадки или в высококормных озерах- 1700 г. Чир и гибрид пеляди с чиром превосходит по темпу роста пелядь на 20-30 %. Муксун в первые два года имеет примерно такой же темп роста, как пелядь, в дальнейшем превосходит ее. Карп в двухлетнем возрасте достигает к октябрю массы 450-500 г. Двухлетки белого амура достигают массы 150-250 г, пестрого толстолобика-350-430 г, белого толстолобика-100-150 г. Товарную рыбу вылавливают из озер закидными неводами, ставными сетями, ловушками, ставными неводами, тралами и плавными сетями. IX этап. Вылупление (220 сут). Зародыши освобождаются из оболочек не все одновременно. Выклев начинается с появления единичных личинок, затем количество их увеличивается. Момент, когда в аппарате уже имеется несколько десятков личинок, можно считать началом выклева. При хороших условиях развития число личинок быстро увеличивается, и наступает массовый выклев. Освобождение зародышей от оболочек происходит следующим образом. В железе вылупления зародыша образуется особое вещество, которое с приближением стадии выклева начинает выделяться из железы вылупления в перивителлиновое пространство. Выделение фермента происходит постепенно и приводит к перевариванию оболочек, в результате снижается их прочность. Двигаясь в ослабленных оболочках, зародыш растягивает, а затем и разрывает их. Сначала появляется голова личинки или хвост, а потом освобождается вся личинка. Свободный эмбрион. Запасы желтка сильно сокращены. Печень позади желточного мешка. Грудные плавники сильно увеличены
Таблица 6. Эмбриональный период развития омуля
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Последнее изменение этой страницы: 2019-03-30; Просмотров: 335; Нарушение авторского права страницы