|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Тема 15. ЦИТОГЕНЕТИЧЕСКИЙ МЕТОД
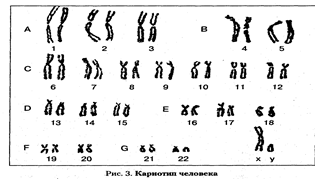
Основа метода — микроскопическое изучение хромосом человека. Цитогенетические исследования стали широко использоваться с начала 20-х гг. XX в. для изучения морфологии и подсчета хромосом человека, культивирования лейкоцитов для получения метафазных пластинок. Развитие современной цитогенетики человека связано с именами цитологов Д. Тио и А. Левана. В 1956 г. они первыми установили, что у человека 46, а не 48, как думали раньше, хромосом. Это событие положило начало широкому изучению митотических и мейотических хромосом человека. В 1959 г. французские ученые Д. Лежен, Р. Тюрпен и М. Готье установили хромосомную природу болезни Дауна. В последующие годы были описаны многие часто встречающиеся у человека хромосомные болезни. Цитогенетика стала важнейшим разделом практической медицины. В настоящее время цитогенетический метод применяется для диагностики хромосомных болезней, составления генетических карт хромосом, изучения мутационного процесса и др. В I960 г. в Денвере (США) была разработана первая Международная классификация хромосом человека. В ее основу легли размеры хромосом и положение первичной перетяжки — центромеры. Все хромосомы по форме разделены на метацентрические, субмета-центрические и акроцентрические и подразделены на 7 групп, обозначенных латинскими буквами А, В, С, D, Е, F и G. Каждая пара хромосом обозначена порядковым номером от 1 до 23, отдельно выделены половые хромосомы — Х и Y (рис.1) У женщин две Х-хромосомы, у мужчин Х и Y-хромосомы. Х-хромосома у женщин не отличается от аутосом группы С; Y-хромосома акроцентрическая, сходная с хромосомами группы G, не имеет спутников. Длина короткого плеча может значительно изменяться. Аутосомы группы С и D содержат в коротких плечах районы ядрышкового организатора.
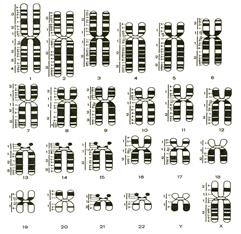
В 1971 г. на I Пражской конференции генетиков в дополнении к Денверской классификации были представлены методы дифференциальной окраски хромосом, благодаря которым каждая хромосома приобретает свой неповторимый рисунок, что помогает точной идентификации. Основные сведения о морфологии хромосом человека получены при изучении их в метафазах митоза и профазе-метафазе мёйоза. При этом важно, чтобы количество делящихся клеток было достаточно высоко. Важнейшие цитогенетические работы выполнены на лимфоцитах периферической крови, поскольку культивирование лимфоцитов в течение 2—3 суток в присутствии фитогемагглютинина позволяет получить множество метафазных пластинок для хромосомного анализа. Цитогенетическому анализу подвергают однослойные метафазные пластинки с раздельно лежащими хромосомами. Для этого делящиеся клетки обрабатывают колхицином и некоторыми другими химическими веществами (гипотоническим солевым раствором, метанол-уксусным фиксатором и др.). Важным этапом цитогенетического анализа является окраска полученных препаратов. Ее проводят простыми, дифференциальными и флюоресцентными методами. Простая окраска обеспечивает групповую идентификацию хромосом. Используется она для количественного учета хромосомных аномалий при определении мутагенности среды (действия радиации, химических мутагенов и др.). С помощью этого типа окраски были открыты многие хромосомные болезни, а также хромосомные аберрации, вызывающие самопроизвольные аборты, врожденные пороки развития, канцерогенез и т. п. В 70-е гг. XX в. в медицинской практике начали применяться методы дифференциального окрашивания, выявляющие структурную разнородность хромосом по длине, что выражается в виде чередования светлых и темных полос (эу- и гетерохроматических районов). Отмечается, что протяженность и рисунок полос специфичны для каждой хромосомы.
Дифференциальное окрашивание хромосом можно проводить рядом способов. Первоначально использовали акрихин-иприт-флюоресцентное алкилирующее вещество (Q-метод). Действие его основано на способности метафазных хромосом дифференциально связывать флюорохромы. После окрашивания акрихином сегменты приобретают яркое флюоресцирующее свечение. Рисунок каждой хромосомы специфичен по числу, размерам и положению по-разному флюоресцирующих сегментов, что и обеспечивает идентификацию всех хромосом. С помощью данного метода окраски можно идентифицировать хроматин с повышенным содержанием АТ-пар, поскольку они активнее флюоресцируют. Специфическим преимуществом Q-метода является то, что он позволяет даже в интерфазном ядре идентифицировать Y-хромосому по яркому свечению. Для просмотра таких препаратов используют люминесцентный микроскоп. В дальнейшем был разработан способ окраски хромосом без флюоресцентных красителей — G-окраска (краситель Гимза). После предварительной инкубации в солевом растворе хромосомы обрабатываются протеазой. В результате хромосомы приобретают сегментированный вид благодаря чередованию темно- и светлоокрашенных участков. Механизм образования сегментов пока недостаточно ясен. Предполагается, что окрашенные сегменты - это гетерохроматиновые участки с повторяющимися последовательностями ДНК, а неокрашенные — это эухроматиновые районы с кодирующими последовательностями ДНК. К разновидностям дифференциального окрашивания по методу Гимзы относятся R-окрашиваемость и С-окрашиваемость. Эти разновидности дифференциального окрашивания получают при определенном изменении времени и условий инкубации препаратов, окрашенных по методу Гимзы. В первом случае распределение окрашенных и неокрашенных сегментов будет обратным тому, что наблюдается при G- и Q-окрашивании. На R-окрашенных хромосомах гетерохроматиновые районы (центромерные, околоцент-ромерные и интерстициальные) остаются светлыми. В случае же С-окраски выявляются районы структурного или факультативного гетерохроматина. В хромосомах человека эти районы локализованы в околоцентромерных участках, а в Y-хромосоме — в дистальной половине длинного плеча. Наиболее крупные блоки С-хроматина имеются в области вторичных перетяжек аутосом 1,9 и 16, а также в Y-хромосоме. Самыми мелкими центромерными блоками обладают Y-хромосома и аутосома 2. Одной из особенностей хромосом человека является асинхронность (неодновременность) репликации по длине. В каждой хромосоме есть рано и поздно реплицирующиеся участки. Для выявления последовательности репликации применяется 5-бромдезоксиуридин — аналог тимина. Включившие его участки окрашиваются слабо. Применяется 5-бром-дезоксиуридин и для дифференциальной окраски сестринских хроматид, если он вводится на полный клеточный цикл. В этом случае вновь образуемая хроматида включит этот аналог тимина и будет окрашена слабо, а другая (старая) окрасится интенсивно. Этот метод позволяет выявлять участки сестринских хроматидных обменов (СХО). При воздействии различными мутагенными факторами число СХО увеличивается, следовательно, этот метод выгоден для изучения мутационного процесса у человека. Успехи молекулярной цитогенетики человека позволяют разрабатывать новые методы изучения хромосом. Так, следует отметить метод флюоресцентной гибридизации in situ (FISH-метод), который дает возможность исследовать широкий круг вопросов от локализации гена до расшифровки сложных перестроек между несколькими хромосомами. Метод FISH может применяться и для диагностики анеуплоидий в интерфазных ядрах. Таким образом, соединение цитогенетических и молекулярно-генетических методов в генетике человека делает почти неограниченными возможности диагностики хромосомных аномалий.
|
Последнее изменение этой страницы: 2019-03-31; Просмотров: 252; Нарушение авторского права страницы