|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
В непрямом конкурентном формате ИФА используются меченные ферментом антитела (специфические или вторичные) и иммобилизованный на твердой фазе конъюгат антиген-белок-носитель.
Непрямая схема с использованием меченых антивидовых антител является одной из наиболее распространенных схем ИФА. На поверхности носителя иммобилизуют конъюгат антиген-белок, к которому добавляют раствор, содержащий определяемый антиген и фиксированную концентрацию немеченых специфических антител, инкубируют и после удаления несвязавшихся компонентов добавляют фиксированную концентрацию меченых антивидовых антител. После инкубации и отмывки носителя детектируют ферментативную активность образовавшихся на твердой фазе специфических иммунных комплексов, причем величина сигнала находится в обратно-пропорциональной зависимости от концентрации определяемого антигена. Применение универсального реагента — меченых антивидовых антител — даёт возможность выявлять антитела к разным антигенам. Кроме того, анализируемый образец и меченый реагент вводятся в систему на разных стадиях, что устраняет влияние различных эффекторов, содержащихся в образце, на каталитические свойства ферментной метки. Однако такая схема анализа усложняет его проведение из-за введения дополнительных стадий. В зависимости от того, какие антигены используются, иммуноферментные тест-системы подразделяют на: Лизатные — в которых используется смесь нативных антигенов (лизированный или обработанный ультразвуком возбудитель инфекции, полученный в культуре); Рекомбинантные — в которых используются полученные генно-инженерным способом белки-аналоги определённых белковых антигенов возбудителя; Пептидные — использующие химически синтезированные фрагменты белков. ОБМЕН И ФУНКЦИИ НУКЛЕОТИДОВ
(16) Биосинтез и катаболизм пуриновых нуклеотидов. Регуляция биосинтеза. De novo (90%)/из готовых пуриновых осн. Источники атомов пуринового кольца (ТГФК – ТетраГидроФолат, он же Н4-фолат):
Фолиевая (птероилглутаминовая) к-та. Производные Н4-фолата – доноры одноуглеродных фрагментов.
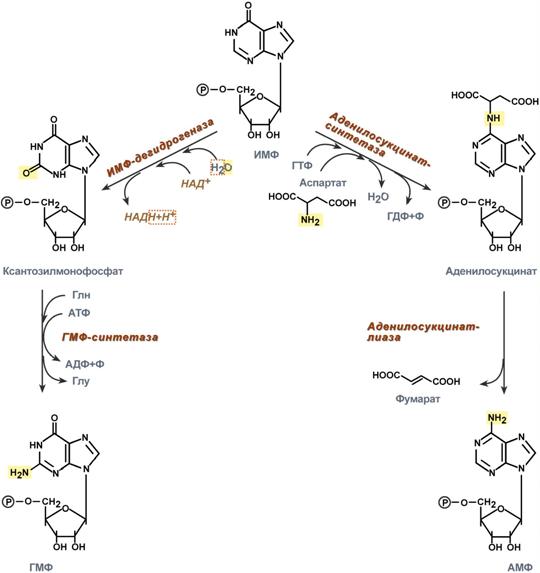
I . De novo : 1) Рибозо-5-фосфат (ФРПФ-синтетаза, ATP → AMP )→ 5-ФосфоРибозил-1-ПироФосфат (ФРПФ, он же ФРДФ) 2) ФРПФ (ФРПФ амидотрансфераза, Gln → Glu ) → 5-ФР-1-амин 3) 5-ФР-1-амин → 7 р-ий, 5 АТР → ИМФ (инозин-5’-монофосфат) Подробнее: ФР-амин NH2-Rib(P) (+ Gly ; ATP → ADP + Pi ) → ГлицинАмид-РибоНклтд NH2-CH2-C(O)-NH-Rib(P) (метенил-Н4-фолат → Н4-фолат) → ФормилГлицинАмид-РибоНклтд CHO-NH-CH2-C(O)-NH-Rib(P) ( Gln → Glu ; 2 ATP →2( ADP + Pi )) → АминоИмадазол-РибоНклтд, цикл замкнулся (+ Asp ; + CO 2; 2 ATP →2( ADP + Pi )) → АминоИмидазол-N-Сукцино-КарбоксАмид-РибоНклтд, к углероду через –С(О)- навесился Asp COOH-CH2-CH(COOH)-NH-C(O)-АминоИмидазол-РибоНклтд (–фумарат) → АминоИмидазол-КарбоксАмид-РибоНклтд (формил-Н4-фолат → Н4-фолат) → ФормАминоИмидазол-КарбоксАмид-РибоНклтд (–Н2О) → Инозин-5’-моноФосфат-РибоНклтд 4) ИМФ → АМФ и ГМФ Подробнее: 1. IMP (ИМФ-дегидрогеназа; + H2 O; NAD+→ NADH+ H+) → КсантозинМоноФосфат (ГМФ-синтетаза; Gln→ Glu; ATP→ ADP+ Pi) → GMP: для синт. GMP требуется 7 ATP 2. IMP (аденилосукцинат синтетаза; Asp→ H2 O; GTP→ GDP+ Pi) → АденилоЯнтарная к-та (аденилосукцинат лиаза; –фумарат) → AMP: для синт. AMP требуется 6 ATP и 1 GTP 5) Образование НуклеозидТриФосфатов из НМФ: GMP (нуклеозидмонофосфат киназа; ATP→ ADP) → GDP (нуклеозиддифосфат киназа; ATP→ ADP) → GTP AMP (нуклеозидмонофосфат киназа; ATP→ ADP, ADP – в след. р-ию) → ADP (АТР-синтаза; + ADP) → ATP
II . Путь спасения (синтез из аденина и гуанина) Аденин + ФРПФ (Аденин-ФосфоРибозил трансфераза) → АМФ Гуанин/Гипоксантин + ФРПФ (ГГФРТ, Гипоксантин-Гуанин-ФосфоРибозил трансфераза) → ГМФ/ИМФ Регул. синтеза пуриновых нклтд: 3 осн. пути регуляции. (1) ГТФ требуется в синт. АМФ, АТФ – в синт. ГМФ, след. поддерживается баланс А и Г нклтд. (2) ФРПФ-амидотрансфераза – Ключ. ферм., имеет 2 центра аллостер. регул. (для АМФ; для ГМФ и ИМФ). (3) АМФ и ГМФ регул. ФРПФ-синтетазу. Аналоги пуриновых осн. как ингибиторы синт. пуриновых нклтд (противоопухолевые препараты): 6-меркаптопурин и тиогуанин.
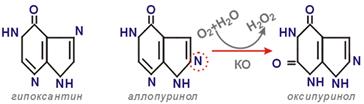
(17) Катаболизм пуриновых нуклеотидов. Причины формирования гиперурикемии. Пути лечебного воздействия при подагре. Катаболизм пуриновых нклтд: 1) Дефосфорилирование АМФ/ГМФ (5'-нуклеотидаза) → Аденозин/Гуанозин 2) Удал. Риб-1-ф. (ПуринНуклеозид фосфорилаза)→ Аденин/Гуанин 3) Удал. NH3 (дезаминаза) → Гипоксантин из АМФ / Ксантин из ГМФ 4) Оксление О2: Гипоксантин (ксантиноксидаза; О2+Н2О→Н2О2) → Ксантин 5) Ксантин (ксантиноксидаза; О2+Н2О→Н2О2)→ Мочевая к-та (0,5-1 г/сут, выводится с мочой) Ксантиноксидаза сод. Мо2+ в кач-ве кофактора.
Гиперурикемия. В сыв. N до 70 мг/мл (0,42 ммоль/л) (чуть выше предела растворимости мочевой к-ты). Min урикемия → обр. кристаллов уратов. Причины: (1) наследств. – повр. двух ферм. (а) ↑акт. ФРПФ-синтетазы (полиморфизм гена) (б) дефиц. ГГФРТ. (2) обильный прием пищи, богатой пуринами (телятина, печень, благородная рыба, икра, кофе, шоколад). (3) повыш. обр. и распад пуринов при гематологич. заб. (обр. и расп. много ядерных кл.): лейкоз, миелоидная метаплазия, расп. злокач. кл. в рез-те лечения цитостатиками. Заб., связ. с метаб. пуриновых нклтд: (1) подагра. Ураты откл. в суст. хрящах, синовиальн. обол., подкожн. клетч. Периодич. – приступы остр. восп. Нефропатия. Причины обычно генетич. Лечение: Аллопуринол (структ. аналог гипоксантина) → Оксипуринол (конкур. ингибитор). Т.о., накаплив. гипоксантин (*10 раз лучше раств., чем урат). Аллапуринол вз/д с ФРПФ в пути спасения, обр. нуклеотид, кот. ингиб. ФРПФ-синтетазу и ФРПФ-амидотрансферазу → торм. синтез de novo . (2) синдром Лёша-Нихана. Тяж. форма. Подагрические узлы, нефропатия, повр. НС. Без лечения – смерть до 10 лет. Рецесс. призн. в Х-хр-ме. Отсут. ГГФРТ (осн. ферм. пути спас.) → Гипоксантин и Гуанин не исп. в пусти спас., полностью превр. в мочевую к-ту → синт. пуринов de novo → ↑распад. Лечение: Аллопуринол, но неврологич. повр. не излеч., т.к. гипоксантин токсичен для мозга. (3) ксантурия. Нед-ть ксантиноксидазы. Накапл. гипоксантин. (4) дефицит аденозиндезаминазы и пуриннуклеозидфосфорилазы → накапл. дезоксиНТФ → инглиб. синт. ДНК в лейкоцитах.
(18) Биосинтез и катаболизм пиримидиновых оснований. Регуляция биосинтеза. I . De novo. Синт. кольца, затем присоед. его к ФосфоРибозилПироФосфату.
(1) ключ. р-ия, ферм. КарбамоилФосфатСинтетаза II (2 – в ц/пл, 1 – только в мтх гптц, S=NH3) Gln + CO2 + H2O + 2ATP → H2N-C(O)-OPO3H2 + Glu + 2ADP + Pi (2) H2N-C(O)-OPO3H2 + Asp ( Карбамоил - Аспартат трансфераза ) → H2N-C(O)-NH-CH(COOH)-CH2-COOH + Pi (3) H2N-C(O)-NH-CH(COOH)-CH2-COOH (ДеГидроОротаза) → дигидрооротат + Н2О (4) дигидрооротат ( дигидрооротат дегидрогеназа ; NAD+→NADH+H+) → оротат (5) оротат ( Оротат - Фосфорибозил трансфераза ; ФРПФ →PPi) → оротдинмонофосфат (6) оротдинмонофосфат (ДеКарбоксилаза оротдин-5’-монофосфата) → УридинМоноФосфат + СО2 Синтез УТФ из УМФ: UMP + ATP (НМФ-киназа) → UDP + ADP UDP + ATP (НДФ-киназа) → UTP + ADP ЦТФ синт. из УТФ: UTP + Gln + ATP (ЦТФ-синтетаза) → CTP + Glu + ADP + Pi
II . Путь спасения. 1) Урацил + Rib-1-P (нуклеозид фосфорилаза) → Уридин + Pi 2) Уридин + ATP (уридин киназа) → UMP + ADP Регуляция синтеза. Рег. – КАД-фермент (полифункц. фермент: КФС II, аспартаттранскарбамоилаза и дигидрооротаза). UMP и др. нклтд аллостерически ингибируют КАД-фермент, ФРПФ – активирует. Карбамоилфосфатсинтетаза II – ключ. фермент. Катаболизм. Цитидин (–NH3) → Уридин (–Rib-1-P) → Урацил (NADPH+H→NADP+) → дигидроурацил (+Н2О) → β-УреидоПропионовая к-та (+Н2О) → NH3 + CO2 + NH2-CH2-CH2-COOH Т.о., Цитидин и Уридин – в β-Ала, Тимидин – в β-аминоизобутират.
(19) Синтез пиримидинов de novo. Причины формирования оротацидурии. Пути лечебного воздействия при оротацидурии. Оротацидурия. Причины: (1) нед. ферментов образования и декарбоксилирования оротидин-5’-Ф (не обр. УМФ) → накапл. оротовая к-та. Пиримидиновый голод, гибель в первые годы жизни. Лечение: вв. УМФ. (2) гипераммониемия. Наруш. любого, кроме первого, ферм. синтеза мочевины. Карбамоилфосфат из мтх → в ц/пл → изб. на синтез пиримидинов. (3) лечение аллопуринолом. Ингиб. декарбоксилазу оротидинмонофосфата (6) → накапл. оротат. Биосинтез дезоксирибонуклеотидов. Акт. только на стадиях подготовки к синт. DNA. Осущ. мультиферм. комплекс – РибоНуклеотидРедуктаза (РибоНуклеозидДифосфатРедуктазный комплекс). Субстр. – дифосфаты. ADP (РибоНуклеозидДифосфатРедуктаза; тиоредоксин-( SH )2→тиоредоксин< SS + H 2 O ) → dADP (киназа; ATP → ADP ) → dATP GDP → dGDP → dGTP; CDP → dCDP → dCTP. Обратно - NAD-dep. (тиоредоксин восст.) Для dTTP: Действие противоопухолевых преп. в синт. пиримидинов: метотрексат (аналог фолата, ингиб. дигидрофолатредуктазу), 5- F-урацил (синт. F-dUMP, ингиб. тимидилат синтазу), 5- F- UMP → 5- F- UTP (встр. в RNA, кот. утрач. норм. ф-ии).
(20) Строение и физико-химические свойства ДНК. DNA – полинклтд. Перв. стр-ра – цепь мононклтд, соед. фосфодиэфирной св. Втор. – двуцепочечная стр-ра. Направление антипараллельное. Удерж. с пом. водор. св. А=Т, Г≡Ц. Также удерж гидрофобн. св. (стекинг-вз/д). B - DNA 1 виток=10 пар нклтд, d2нм, l=1,5-1,7 нм. A - DNA виток шире, короче (11 пар), хар-на для репликации. Z - DNA 9 пар. Стадия интерфазы. 2sDNA кольц. – прокариотич., 1sDNA кольц. – у бактериофага. Трет. стр-ра – DNP. Стр-ра нуклеосом, d=11 нм. Нуклеосома сод. ок. 200 пар нклтд, имеет кор (сердцевину). Белки (гистоны, (+)-зар.), фосфатн. гр. → удерж. 8 мол. гистонов составляют кор, вокруг которого – 1,75 витка DNA. Линкерная DNA – ок. 50 пар нклтд, присутств. гистон Н1. Вокруг кора – 146 пар нклтд + 50 пар линкерной DNA. Четв. стр-ра. Цепочка укорач., утолщ., d=130 нм. Формирование петель, удерживаемых негистоновыми белками (хромомеры). Обр. хромонема → хромосома (d=1400 нм). мтхDNA – циклич. стр-ра, 16569 пар нклтд, кодир. 13 б. (б. дых. цепи), кодир. tRNA (22 гена). 20% хроматина – негистон. б. Все нигстон. б. имеют четкие сайты связ. с DNA (цинковые пальцы). Н/г б.: 1) цинковые пальцы (5 сайтов св.), 2) HMG-белки (оч. высок. скорость движ. при ЭФ), 3) б., в сост. кот. есть втор. надстр-ра (спираль-поворот-спираль). Физ-хим св-ва: М 3*10^9 Да. Показатель плотности: метод ультрацентриф. (ЦФ зонально-скоростное, CsCl), =1,69-1,73. Есл денатурир., то плотность выше. Денатурация DNA – разруш. неков. водор. св. Факторы: тем-ра, рН, мочевина в опр. концентр. Плавление DNA: 50-60˚ денат. нет, при ↑тем-ры проявл. гиперхромный эфф., далее – эфф. кооперативности. Тплав = тем-ра, при кот. 50% DNA денатур. (у чел-ка – 81,5˚, у E. coli – 90,5˚). Тплав зависит от кол-ва пар G≡C. Отжиг – медленн. охл. с возм. формирования обратно 2sDNA.
(21) Методы исследования структуры ДНК. Гибридизация, секвенирование, ПЦР. Рестриктазы – бакт. эндонуклеазы, разрез. мол. DNA . 3 типа: 1) без обр. компл. фрагм., 2) с обр. липких концов, 3) несимметр. (не палиндромы). Сейчас исп. 600+ разл. рестр., чаще – класс 2. (1) (а) Метод гибридизации. 1969. Плавление 2 разн. фрагм. DNA, обр. денатурир. спирали, от кажд. забир. по 1s → отжиг. Чем больше компл. уч-ков, тем быстрее. Примен. гибр.: док-во родства, картирование генов у прокариот, опр. перв. стр-ры. (б) Метод блот-гибридизации (Саузерн-блоттинг). Бл. – перенос фрагм., разд. методом гелевого ЭФ, на тверд. носит. (нейлон, нитроцеллюлоза) под действ. капиллярных сил. Саузерн: исп. радиоакт. зондов. Внос. доп. краситель, далее – присоед. овидин. Внос. щелоч. фосфатаза и ее S (изм. ск. абс.). Либо: флуоресц. метки (не нужно прокраш.) = FISH. Исп. зондов при диагн. генетич. заб. (in situ: можно опр. один ген). (2) Секвенирование. Опр. перв. стр-ру. (а) Сенджер. 800-1000 нклтд. ДиДезоксиСекв. Синт. изучаемой DNA in vitro с остановкой синт. путем присоед. dd-нклтд, у кот. отсутств. 3’-OH → «метод обрыва цепи». dd -нклтд = терминаторы. Матрица – 1sDNA, к кот. присоед. радиоакт. меченый праймер. 4 пробирки (матрица DNA , праймер, DNA - pol ., 4 dNTP , буфер с Mg 2+) → в каждую пробирку – по одному dd -нклтд → гель, 4 лунки, эл. ток → разделение с разрешением в 1 нклтд → опр. посл-ти. Усовершенствование метода: замена радиоакт. метки на флуоресц. 2001 – расшифр. геном чел-ка с пом. секв. (б) Пиросеквенирование. 1996. Синт. DNA DNA-pol., сопр. с работой ферм. люциферазы (хемилюминесц. метод). Рег. присоед. кажд. нклтд., высвоб. пирофосфат PPi. Ферм. сульфурилаза → встр. а ATP, ферм. люцифераза превр. люциферин в оксилюцеферин → свет. Метод компьютеризирован. Нклтд вводятся кадж. отдельно, присоед. через пористый планшет. Сейчас – ок. 1000 нклтд. (3) ПЦР. Компоненты: DNA-матрица, сод. тот уч-ок, кот. треб. амплифиц.; 2 праймера; Термостаб. DNA-pol. (Taq-пол., Pfu-пол., Pwo-пол.), dNTP, Mg2+, буфер. +Масло, чтобы смесь не испарялась. Ход: обычно 25-30 циклов. 1) денатурация, 2) отжиг, 3) элонгация. (4) Клонирование. Вв. вектор в бактерию/бактериофага.
(22) Строение и функции РНК. tRNA, mRNA, rRNA; scRNA (small cytoplasmic), snRNA (small nuclear) SNUPRS = Small NUclear Ribonucleoprotein ParticleS mRNA: инф. о посл-ти а/к в б. tRNA – малые, ок. 80 нклтд, с консерв. третичн. стр-рой. Кажд. tRNA сод. уч-ок для присоед. а/к и антикодон для узнав. и присоед. к кодонам mRNA. Антикодон обр. водор. св. с кодоном → tRNA в положение для обр. пепт. св. между последней а/к образ. пептида и а/к, присоед. к tRNA. rRNA – каталитич. сост. рибосом. Эу: 18S, 5.8S, 28S и 5S Ферм., необх. для превращ. НК: (1) Нуклеазы (гидралазы): ДНКазы/РНКазы, Эндо-экзонуклеазы. (2) Лигазы: ДНКлигаза/РНКлигаза. Устран. единичн. разрыв внутри цепи НК. Зашивают ник. (3) Топоизомеразы: 1. ДНК-т/и I разрывает одну из цепей суперспирал. DNA, спираль раскручивается, ДНК-т/и I устраняет разрыв АТР-независимо. В акт. центре – остаток Tyr; 2. ДНК-т/и II разрывает сразу 2 цепи. У про- – гираза. АТР-зав. Проводит сквозь разрыв фрагм. 2sDNA, затем сшивает, участв. в введ. (+) и (–) витков. В акт. ц. – Tyr. (4) Хеликаза. Расплетающ. б. Разрыв. водор. св. между аз. осн. Подд. DNA в распл. сост., продвигает вилку.
МОЛЕКУЛЯРНАЯ БИОЛОГИЯ (матричные биосинтезы) (41) Репликация ДНК у прокариот. Свойства ДНК-полимераз прокариот. Ингибиторы репликации.
Синтез (репликация, удвоение) ДНК – в S-фазу кл. цикла. Мех-м репликации: Мезельсон и Сталь в 1957 г, полуконсерв., т.е. на кажд. нити матер. ДНК синтез. дочерняя копия. Компоненты репл.: матрица, растущая цепь, субстраты для синтеза (dNTP), источник энергии, ферменты. У Про-: инициация, элогнация, терминация. У E. coli OriC – 240 пар осн. Ori=origin (uni-/bidirectional). Топоизомераза I/II, осущ. репл., не узнает Origin. (1) Инициация. dnaA узнает OriC (АТР-зав.). Деспирализация – после работы Топоизомеразы. Новая цепи строится 5’→3’. Хеликаза у E. coli – dnaB. SSB-белки (Single Strand DNA-Binding, защищ. ее от эндонуклеаз, не дают обр. 2sDNA), в каждой вилке – 200+ SSB. При нач. реплик. освоб. нить. Праймаза (RNA-pol.) создает праймер, работает 1 раз на лидир. цепи, на отстающей – для кажд. фрагм. Оказаки. Праймер – неб. уч-ок RNA (4-12 нклтд), компл. родительской 1sDNA. Обеспеч. наличие ОН-гр. в 3’ Rib. Праймосома – комплекс всех факторов репликации (все перечисленное, кроме dnaA и Топ/изом I/II). Ф-ии праймосомы: нач. синт. DNA около Ori, движется за счет эн. АТР, создеат праймеры. (2) Элонгацию осущ. DNA- pol. DNA- pol. III – ведущая, раб. 5’→3’, катализ. синт. ведущ. и ост. цепи, узнает фрагм. Оказаки. Нач. синт. с праймера. Акт в 50 раз ↑ DNA-pol. I, в 1000 раз ↑ DNA-pol. II. М=422 кДа. 10 суб/ед трех типов (альфа, эпсилон, тета). Эпсилон – корректирующая акт-ть (3’→5’ экзонуклеазная акт-ть), альфа – полимериз. акт-ть, тета – сохран. конформ. Ск. синт =30 000 нклтд/мин. DNA- pol. I. М 143 кДа. 3 акт-ти: 1) полимеризационная (удлин. фрагм. Оказаки 5’→3’), 2) корректирующ. (3’→5’ экзонуклеазная акт-ть), 3) репликационн. (5’→3’ экзонуклеазная акт-ть). Вырез. 1 RNA-нклтд, сзади дострив. DNA-нклтд (мех-м удаления RNA-праймера) → остается nick, кот. зашивается лигазой. Ск. синт =600 нклтд/мин. DNA- pol. II. М=90 Да. 1 цепочка. Предп., необх. для репарации. 3’→5’корректир. акт-ть. Ск. синт =30 нклтд/мин. (3) Терминация. TER-посл-ть сост. из 23 пар нклтд. Их узнает TER-binding protein (преп. повт. раскруч. DNA, останавл. работу праймосомы). Процесс репл. осущ. макромолекулярным комплексом – РЕПЛИСОМОЙ (часто присоед. к ядерн. мембр. изнутри). Ингибиторы репл. у про-: дауномицин и доксрубицин (интеркалируют между G≡C в мол. DNA), актиномицин D (блок. синт. DNA и RNA у про- и эу-), алкилирующие агенты и ингибиторы ДНК-топоизомеразы II (ингибиторы гираз).
(42) Репликация ДНК у эукариот. Свойства ДНК-полимераз эукариот. Ингибиторы репликации. См. 41, НО: В синт. Эу- DNA уч. 5 DNA - pol: α , β , γ , δ , ε. α,δ – на лид. цепи, α,δ/ε,β – на отстающей. α (I) иниц. репелик. (синт. праймера: 8-10 Rib-нклтд)+ ок. 50 dRib-нклтд, β (III) устраняет бреши между фрагм. Оказ., γ – мтх DNA, δ (II) и ε: RNA-праймер на ост. цепи удлин. до встречи с др. праймером.
(43) Инициация, элонгация и терминация транскрипции у прокариот. ДНК- зависимая РНК-полимераза прокариот. Трнскр – счит. инф. с 1sDNA на мол. mRNA. У про- сопр. с трансляцией, обр. готов. mRNA; rRNA и tRNA обр. в виде предшеств. Факторы трнскр: DNA - d RNA - pol (только полимеразная акт-ть 5’→3’), DNA -матрица, NTP, Mg 2+ (катализ). DNA - d RNA - pol у про-: 5 с/ед (2α – 40 кДа, форм. каркас RNA-pol, спос. вз/д с промотером; β – 155 кДа, RNA-pol акт-ть; β’ – 160 кДа, св. ферм. с DNA, есть шип; ω – 60 кДа, то же, что α). + σ (85 кДа) – иниц. транскр. (м.б. много). У компл. бол. (–) зар. (наиб. – у σ) – нар.; внутр. – (+) зар. → хор. св. с фосф. гр. DNA. В акт. ц – Mg2+ (для присоед. нклтд). NTP св. с Mg2+ → необх. 2 иона. В β’ – шип (разделяет DNA-RNA гибрид). Ед. трнскр = оперон. p–o–A–B–C–t. p → узнав., инициация. o → оператор (св. индукт. или репресс., вкл./выкл. p + o = 5’- UTR. t=terminator=3’- UTR (UnTranslated Region). A, B, C – структурные гены. Интронов нет. → ПОЛИЦИСТРОННАЯ mRNA (код. неск. б., у эу- – в мтх). Ингибиторы транскрипции: рифампицины (ингиб. бакт. DNA-d RNA-pol, препятств. инициации транскрипции), интеркаляторы (актиномицин D, антималярийные преп.), гепарин (преп. св. с промотером), амантин (яд блед. пог., блок. акт-ть RNA-pol III).
Этапы. (1) Инициация. Праймер не треб., т.к. есть промотор, кот. нацелив. DNA-d RNA-pol. Присоед. DNA - d RNA - pol → start point (где присоед. 1 нклтд, далее → 3’=upstream=(–), →5’=downstream=(+) ). TATAAT = Pribnow box, часть промотора (энхансер), прибл. на 10 пар нклтд выше точки начала трнскр. Ведущая с/ед у про- в иниц. – σ (присоед. к ТАТААТ). В обл. upstream (→3’) – промотор (узнав. по 2 точкам). Энхансеры – акт., сайленсеры – торм. (2) Элонгация. Рег. мол. действ. на факт. транскр. (TBP-б. → рег. генов, отв. за p53, NFκβ, эстрогенов. Rs). Синт. pre-RNA. У про- и эу- одинаково. RNA-pol 5’→3’: узнав. промтороа, плавление промотора, движ. фермента, синтез, до ter. (3) Терминация. 1) при участии ρ-фактора (гексамер): св. с растущей цепью RNA, движ. → догоняет RNA-pol и св. с ней; на DNA – ter уч-ок (мн. C, G), б. св. с ферм., ведет к расплет.; ρ св. с RNA-pol. 2) без ρ-белка. В обл. ter – палиндромы (мн. C, G), далее – мн. А («шпилька»).
(44) Инициация, элонгация и терминация транскрипции у эукариот. ДНК- зависимые РНК-полимеразы эукариот. DNA - d RNA - pol у эу-: 3 ядерных, 1 мтх. Гетеромультимерн. компл. Сходство с про- в β-с/ед (символика другая). RNA-pol I – 14 с/ед – ядрышки – 45S rRNA RNA-pol II – 12 с/ед – нукл/пл – hnRNA, snRNA RNA-pol III – 17 с/ед – нукл/пл – tRNA, 5S rRNA, scRNA, snRNA Ед. трнскр = транскриптон. Зоны: информативная (стр. гены: интроны и экзоны, терминатор) и неинформативная (промтор, гены оператора). → МОНОЦИСТРОННАЯ mRNA Ингибиторы транскрипции: рифампицины (ингиб. бакт. DNA-d RNA-pol, препятств. инициации транскрипции), интеркаляторы (актиномицин D, антималярийные преп.), гепарин (преп. св. с промотером), амантин (яд блед. пог., блок. акт-ть RNA-pol III).
Этапы. (1) Инициация. Праймер не треб., т.к. есть промотор, кот. нацелив. DNA-d RNA-pol. Присоед. DNA - d RNA - pol → start point (где присоед. 1 нклтд, далее → 3’=upstream=(–), →5’=downstream=(+) ). У эу- в промоторе – ТАТА-box = бокс Голдберга-Хогнесса. RNA-pol I – промотор аналогичен прокариотическому; ТАТА. RNA-pol II – синт. hnRNA; 3 тчк узнав.: TATA box, CAAT box, GC box. RNA-pol III – счит. инф. для 5S, для них промотор в обл. downstream. Энхзанс. – акт., сайленс. – торм. Ок. 10% генома – кодировка факт. транскр. (ок. 2 600 б.) Б.-факт. транскр. сод. DNA-binding домен или комплем. др. факт. транскр. (2) Элонгация. Рег. мол. действ. на факт. транскр. (TBP-б. → рег. генов, отв. за p53, NFκβ, эстрогенов. Rs). Синт. pre-RNA. У про- и эу- одинаково. RNA-pol 5’→3’: узнав. промтороа, плавление промотора, движ. фермента, синтез, до ter. (3) Терминация. У эу- есть факт. терминации. ter сод. мн. А. RNA-pol дефосфорилир. → неакт.
(45) Процессинг РНК у прокариот и эукариот. Процессинг предшественников RNA (посттрнскрипционная модификация) про-: mRNA в готовом виде, трнсл по ходу трнскр. pppA, pppG на 5’-конце для защиты. pre-rRNA (E.coli) обр. 30S: 7 дискретн. фрагм., кажд. вкл. копирование; соед. спейсерами, на кот. tRNA. 2 ферм.: RNAase III, RNAase P , напр. проц. от 5’ к 3’. Узнав.: кажд. rRNA форм. «шпильки». Одноврем. с обр. rRNA образ. tRNA: –[16S]–[tRNA]–[23S]–[5S]–. Процессинг tRNA (есть и отд. предш.): кластеры (2-7 tRNA) в pre-tRNA. Ферм.: RNAase P (5’→3’ эндонукл.), RNAase D (экзо 3’ акт., отщепл. по 1 нклтд до фрагм. ССА), нуклеотидилтрансфераза (если нет ССА, она достроит 3’). Для tRNA идет модиф. азот. осн. эу -: pre-rRNA синт. RNA-pol I в ядрышке – 45S (18S+5,8S+28S). + 13 000 нклтд. Процессинг экзонуклеазами. Вырез. спейсеры, перед этим – метилирование отд. фрагм. для защиты. Для 5S – спец. ген, кот. транскр. в нукл/пл RNA - pol III одновр. с трнскр. pre-tRNA. Процессинг pre-tRNA: 1) модиф. азот. осн. 2) ! удаление интрона (14-16 нклтд) сплайсингом. 3) форм. антикодоновой петли. 4) удал. лишн. ф. 5) удал. с 3’-конца лишн. посл., присоед. ССА. Первичн. транскрипт 100 нклтд; затем ок. 80. RNAase P , RNAase D. Процессинг pre-mRNA гетерогенноядерн.; коротк. 500-5000. 1) удал. лишн. посл. с 5’ и 3’-концов pre-mRNA (эндонуклеазы; отр. до нетранскр. уч-ков – инф. о промоторе и терминаторе). 2) ! кэпирование 5’-конца (кэпирование – защита от действ. эндонуклеаз + идет более эфф. трансл. + работа ферментн. сис-мы, обеспеч. удал. интронов, осущ. ферм. гуанилилтрансфераза). 3) встр. полиА хвоста к 5’ (полиА-полимераза синтезирует, полиА-нуклеотидтрансфераза перебрасывает, лигаза пришивает; полиА опр. время жизни mRNA, обеспеч. выход из ядра, замедл. гидролиз в ц/пл). 4) сплайсинг. 5) модиф. опр. аз. осн.
(46) Активирование аминокислот. Инициация трансляции у прокариот и эукариот. Трансляция («перевод») – перевод инф., залож. в посл-ти нклтд mRNA в посл-ть а/к ост. в полипепт. (ПП) цепи. Посл-ть нклтд однозначно опред. расп. а/к ост. в ПП цепи. Трнсл лок. в ц/пл. Нарастание с N-конца к C-концу. Рибосома идет 5’→3’ (полирибосома, компл.). Этапы: 1) Активация а/к и присоед. их к соотв. tRNA. 2) Собств. трансляция – сборка ПП цепи с пом. рибосом. Акт-ия а/к: в ц/пл. Необх. а/к, tRNA, ферм. АминоАцил- tRNA -синтетаза (4 акт. центра, +Н2О для гидролиза), АТР, Mg2+. H2N-CH(R)-COOH + ATP –(Mg2+)→ H2N-CH(R)-C(O)~O-PO3-Rib-A + 2Pi. ( обр . АминоАцилАМФ ) аминоацилAMP + tRNA –(Mg2+)→ аминоацилtRNA + AMP. ( tRNA – к 3’-концу) Рекогниция – узнавание а/к и tRNA. аа- tRNA (оч. точн.) по строению: 1 цепь (100 кДа) для Val, Leu; олигомерные (одинаков.) для Met; олигомерные (разные) для Gly, Trp. Далее расщ. дифосфата (АТР-зав.) Итого: необх. 2 АТР!
Этапы трнсл: (1) Инициация. К mRNA присоед. мал. с/ед рибосомы (2 уч-ка: Р – пептидный, А – аминоацильный), занимает 2 триплета (в Р-центре всегда -AUG- ). В Р-центр → tRNA с Met, компл. соед. с кодоном, потом приходит бол. с/ед. (2) Элонгация. В А-центр с кодоном идет tRNA с соотв. а/к. Раб. пептидилтрансфераза → обр. пепт. св. Транслоказа (+GTP) → перемещ. рибосому по mRNA → в Р-центр переходят связавшиеся а/к. (3) Терминация. В А-центр попадает UGA, UAG, UAA – стоп-кодон. Рибосомальный комплекс разбирается, б. складывается.
(47) Процесс трансляции у прокариот. Ингибиторы трансляции. (1) Инициация: mRNA, tRNA, GTP – 3 факт. инициации. Мал. с/ед. рибосомы так соед. с mRNA, чтобы -AUG- попал в Р-центр. IF-3 → соед. мал. с/ед. GTP-подводящий фактор → поступл. эн. IF-1 ускор. инициацию. mRNA счит. с 5’. посл-ть Shine-Dolgarno компл. 3’-концу 16S rRNA → AUG в Р-центр. С AUG связ. IF-2 (фармилметионин-tRNA). Подходит 50S (бол. с/ед) → 70S комплекс. (2) Элонгация. EF-Tu (GTP-подвод. для кодон-антикодонового узнавания). Встр. след. кодона mRNA в А-центр. EF-Ts (регенерир. мол. GTP для след. шага). Пептидилтрансфераза (в бол. с/ед) переносит а/к от tRNA в Р-центре на а/к tRNA в А-центре → обр. дипептид. EF-G (транслоказа) – ц/пл, имеет ATP-акт-ть, передвиг. рибосому на 1 кодон (20 а/к в с). (3) Терминация. 3 рег. фактора: 1. узнав. UAA, UAG; 2. узнав. UAA, UGA; 3. присоед. к 1 / 2 факт., подв. GTP → разруш. 20 tRNA. Расх. эн. на синт. 1 пепт. св. = 4 макроэргич. св. (2 GTP).
Ингибиторы: тетрациклины (инг. элонгацию, св. с 30S с/ед рибосом и блок. присоед. aa-tRNA в А-центр), левомицетин (присоед. к 50S с/ед рибосом и инг. пептидилтрансферазную акт-ть), β-лактамы (ингиб. ферм., обеспеч. обр. поперечн. св. в стр-ре б. кл.), макролиды – эритромицин (то же, ингиб. транслокацию), аминогликозиды – стрептомицин (инг. инициацию, св. с 30S с/ед, вызыв. ошибки в прочтении mRNA).
(48) Процесс трансляции у эукариот. Ингибиторы трансляции. (1) Инициация. 13 IF. Перв. tRNA → Met. GTP, ATP. Рибосома 40S+60S. eIF-6 св. с мал. с/ед. CBP (cap binding protein) св. вне рибос. с кэп mRNA → спос. св. mRNA с eIF-4A (RNA-хеликаза, расщ. втор. стр-ру) и eIF-4B (мал. с/ед → в прав. полож.; AUG → в А-центр). eIF-1, 3 (ускор. движ.). eIF-2 (GTP- подв.). eIF-4С (факт. ассоциации). eIF-5. (2) Элонгация. То же, что у про-: EF (GTP-подвод. для кодон-антикодонового узнавания). Встр. след. кодона mRNA в А-центр. EF (регенерир. мол. GTP для след. шага). Пептидилтрансфераза (в бол. с/ед) переносит а/к от tRNA в Р-центре на а/к tRNA в А-центре → обр. дипептид. EF-G (транслоказа) – ц/пл, имеет ATP-акт-ть, передвиг. рибосому на 1 кодон. Транспептидаза – далее (эн. не заканч.). EF-2 (цитоплазматич. транслоказа) → передвиж. на 1 кодон. (3) Терминация. Узн. любой из 3 стоп-кодонов (UAA, UAG, UGA). С ним связ. GTPase → распад. 31 tRNA. 31 аа-tRNA-синтетаза в мультиферм. компл.
Ингибиторы: см. 47. Также: вирусы (ингиб. синтез НК и б. хозяина), токсины: α-амантин (инг. эу- RNA-pol), клещевина → рицин (удаляет остаток аденина из 28S rRNA бол. с/ед и инг. синтез б. у эу-), Cor. diphteriae → дифт. энтеротоксин (катализ. ADP-рибозилирование и инакт. факт. элонг. EF-2).
(49) Регуляция экспрессии генов. Про-. Гены: 1) конститутивные (нужны постоянно, в неб кол-вах). 2) индуцируемые. В N транскр. ок. 600-700 б. (конст.). Больш-во генов индуцир. Гены объединены в оперон (уч-ок DNA, кот. сод. инф. о группе функц. вз/св стр. генов и регуляторную зону, контролирующую их транскр.). (1) Индуцируемый оперон. Лактозный оперон E. coli. Индуктор – S (Lac). Гены: А=β-галактозидаза, В=галактозидпермеаза (осущ. трансп. лактозы через пл. мембр.), С=тиогалактозид транспептидаза. R= ген-регулятор (синт. белок, кот. активирует оператор), расп. оч. далеко, обычно upstream. Пост. выраб. репрессор, след. обычно экспрессия этих генов невозм. При появлении индуктора (Lac) репрессор инактивируется → синт. до 50000 мол. β-галактозидазы за неск. с. –5’–R–––p–o–A–B–C–3’– (2) Репрессируемый. Триптофановый оперон. Синт. Trp идет постоянно. Корепрессор – конечн. продукт или др. в-во. R → репрессор (н/акт). Если +корепрессор, то → блок. оператор. Обычно индуцируемыми явл. гены, синт. ферменты катаболич. р-ий, репрессируемыми – анаболич. р-ий. (3) Катаболическая репрессия lac оперона. ↑[Glc] → ↓[cAMP] → ↓Lac (↓акт-ть оперона). БРЦ = белок-рецептор цАМФ (CAP = cAMP-activated protein). CAP+cAMP → связ. с промотором, подстегивает активность оперона. Регуляция экспрессии на уровне трансляции: (1) недостаток tRNA (мн. бактериофаги метаб. tRNA в кл., подавляя синтез б.). (2) ↓акт. аминоацил-tRNAсинтетаз (фаги выр. ингибиторы). (3) ↑ур-нь RNAases, ↑расщепл. RNA (синдром хронической усталости?). (4) фосфорил. факт. инициац. киназами → потеря акт. (5) регул. числом и акт-тью рибосом. Прокариоты: перекрываемость генов (разные рамки считывания).
Эу-. 5 уровней: претранскр., транскр., посттранскр. (процессинга mRNA), трансляц., посттрансляц. (процессинга б.). (1) Претранскр. Отн. к организации генома. Генные сети, высокая пластичность генома, организация зроматина, хим. сост. (2) Транскр. (не хар. прямая субстратная регул.). Транскр. факторы (изв. ок. 3 тыс.), регуляторные посл-ти (энхансеры, сайленсеры, инсуляторы, респонз-элементы). (3) Посттранкрипц. (уровень процессинга mRNA). Альтерн. сплайсинг, альтернат. промоторы, регуляция стабильность mRNA, редактирование mRNA. (4) Трансляц. уровень: то же, что у прокариот. Нед. tRNA, ↓акт. аминоацил-tRNAсинтетаз, ↑ур-нь RNAases, фосфорил. факт. инициац. киназами, регул. числом и акт-тью рибосом. (5) Пострансляц. (процессинга белка). В ц/пл или ER. 5 этапов: фолдинг, гликозилирование, ограниченный протеолиз, стр. модиф. а/к (карбоксилирование), присоед. простетич. гр. (у сл. б.).
(50) Факторы мутагенеза. Виды мутаций. Антимутагенная защита. Мутагенез — внесение изм. в нклтд посл-ть DNA (мутаций). Есть естеств. (спонтанный) и искусств. (индуцированный) м-нез. Естеств. – под возд. на генет. материал мутаг. факт. окр. среды: UV, радиация, хим. мутагены. Мутационная теория: Хуго Де Фриз (Нидерл., 1903), С. И. Коржинский (1899). Положения: 1. Мут. внезапны, как дискр. изм. призн. 2. Новые формы устойчивы. 3. В отл. от насл. изм., мутации не обр. непрер. рядов, не группир. вокруг к.л. среднего типа. 3. Мутации проявл. по-разному и м.б. полезными или вредными. 4. Вер-ть обна. мутаций зав. от числа исслед. особей. 5. Сходн. мутации могут возн. неоднокр. Механизмы: повр. DNA → не репарир. → мутац. Мутац. не проявл., если они произошли на интроне или в экзоне, но вследствие вырожденности генетич. кода не произ. наруш. ТИПЫ (а) по уровню возникновения (1) генные (инверсии, замены (транзиции (Пи-Пи, Пу-Пу), трансверии (Пу-Пи)) – м.б. либо нонсенс – замена смысл. кодона на терминир., либо миссенс – замена кодона, либо сайлент → кодон-синоним; дупликации, инсерции, делеции – ведут к сдвигу рамки считывания (frame shift)). (2) хромосомные (внутрихром. – хр. дупликации, делеции, инверсии, межхром. – транслокации, транспозиции и т.д.). (3) геномные (поли-, анеу-, гаплоидия). (б) по типу аллельных вз/д: (1) рецессивные, (2) доминантные. (в) по хар-ру проявл. в фенотипе по отн к проявл. норм. аллеля: (1) гипо-, (2) а-, (3) анти-, (4) неоморфные. (г) по вл. на жизнеспос.: (1) летальные, (2) вредные, (3) нейтр., (4) полезные. (д) по происх.: (1) спонтанные, (2) индуцибельные. (е) по месту возн.: (1) генеративные, (2) соматические. (ж) по фенотипич. проявл.: (1) биохим., (2) физиол., (3) морфологич. МУТАГЕНЫ: Физические (излучения); Химические; Биологические (вирусы). СВ-ВА МУТАЦИЙ: Спонтанность; Нет порога (предела); Не зависят от продолжительности и интенсивности действия мутагена. Искусств. мутагенез: направленный/ненаправленный. РЕПАРАЦИЯ: исправл. поврежд. Компоненты: (1) DNA-хеликаза — ферм., узн. хим. измен. уч-ки в цепи и осущ. разрыв цепи вблизи от повр; (2) экзонуклеаза — ферм., удал. повр. уч-ок; (3) DNA-полимераза — ферм., синт. соотв. уч-ок цепи DNA взамен удалённого; (4) DNA-лигаза — ферм., замык. последнюю связь в полимерн. цепи.
(51) Молекулярные механизмы канцерогенеза. Пути активации протоонкогенов. В основе лежит дефект генов, ответственных за деление, репарацию или апоптоз. Канцерогенез- молекулярно-генетическая патология, заболевания генетического аппарата. Как и от чего образуются опухоли? Пока точно не известно. Молекулярная онкология –наука, изучающая данную проблему. Дифференциация, рост, наследование признаков описаны не до конца. Злокачественная опухоль - нерегулируемый, автономный, местный тканевой рост. Опухоль- клон или несколько клонов клеток, в которых генетически закреплено свойство неконтролируемого роста. 1)Теория химического канцерогенеза 2)Теория физического канцерогенеза 3)Теория Вирусного канцерогенеза 4) Теория Онтогена
Химический канцерогенез 1775 год – профессиональный рак у трубояистов – опухоль мошонки в результате воздействия хим. веществ. В настоящее время известно около 5 млн. химических канцерогенезов, около 70 тыс. встречаются в течении жизни, около 8 тыс. изучены, около 1,5 тыс. обладают способностью мутагена. Печень способна некоторые вещества переводить в канцерогены. Физические вещества • Фри Кен (изобретатель рентгеновской трубки) постоянно облучал себя и заработал сильнейший рак • Ультрафиолетовое излучение • Гамма – излучение + ионизирующее излучение • Внутриматочные контрацептивы, объекты вводимые в тело человека (пластическая операция)
Вирусный канцерогенез • Группа Papova (Paphilom polilom Vacualization group) • Вирусы герпеса (97% населения) • Гепатитные вирусы • Вирус Энштейна-Бара – лимфома Беркстта – t больше 11 градусов. ( почти в 100% вызывают опухоль) Трансдукция – захват клеточных последовательностей ДНК хозяина вирусной фагой. Src –саркома.
Все позвоночные обладают консервативным геном Src, который родств. к вирусному Src и обладает онкогеном. Src возник в результате трансдукции клеточной Src. Протоонкогены – определенные участки ДНК, способные в результате трансдукции вирусного гена превращаться в вирусные анкогены или после активации в клеточные трансформирующие анкогены. Классификация анкогена: 1. Анкоген – кодирующий протеинкиназу Src. 2. Не тирозиновые raf 3. Анкогены – фосфорил. рецепторов роста 4. Анкогены кодируют факторы апоптоза bcl -2
Пути активации протоокогенеза: • Точечная мутация • Количественный принцип – появление вблизи «молчащего» протоанкогена, активного промотора (экзогенного характера) – вирус или эндогенного. В результате происходит транслокация • Амплификация – увеличение числа копий протоанкогена Гены супрессоры опухолей – гены, производящие продукты, удерживающие опух. рост.
ОБМЕН И ФУНКЦИИ АМИНОКИСЛОТ (52) Источники и пути расходования аминокислот в организме. Азотистый баланс. Реакции образования и детоксикации аммиака в организме. Источники а/к: (1) пищевые белки. 100 г/сут. Все протеолитич. ферм. – гидролазы, пептидазы (КФ3.4), эндо- и экзопептидазы (N- и C-экзо-). Протеолит. ферм. жел. и ПоджЖ выраб. в неакт. форме, необх. ограниченный протеолиз. Биол. полноценные б. встреч. только у животных (молочный б., б. яиц птиц). Злаки дефицитны по Lys, куруруза – по Trp. (2) а/к, обр. при распаде тк. 200 г/сут. Кл. слизистых ЖКТ, ферменты, внутрикл. б. (3) синтез заменимых а/к. 100 г/сут. Предшественники – прод. у/в обмена, орг. к-ты. Осн. р-ия – переаминирование. Синтез заменимых а/к: (1) ПВК –(переаминирование)→ Ала (2) ЩУК –(переаминирование)→ Асп –(амидирование)→ Асн (3) 3-фосфоглицерат → Сер → Цис; Сер ←→ Гли (4) α-КГ –(восстанов. аминир.)→ Глу → Глн; Глу → Про (5) Цикл синт. мочевины → Арг (6) Фен → Тир (7) Glc –[O]→ Сер (углер. скелет Цис) –→ Цис Мет –(вит.В12)→ Hcy –↗ Подробнее: CH3-S-CH2-CH2-CH(NH2)- COOH Met + H4-фолат –(вит.B12)→ 5-метил-Н4-фолат + Hcy SH-CH2-CH2-CH(NH2)- COOH (гомоцистеин) SH-CH2-CH2-CH(NH2)- COOH Hcy + HO-CH2-CH(NH2)-COOH Ser –(цистатионин-синтаза(вит.В6);–Н2О)→ COOH-CH(NH2)-CH2-CH2-S-CH2-CH(NH2)-COOH Цистатионин –(цистатионин-лиаза(вит.В6);+Н2О)→ CH3-CH2-C(O)-COOH + NH3 α-кетобутират + NH3 + Cys SH-CH2-CH(NH2)-COOH Расход а/к: (1) синт. б. (2) катаболизируют до конца (NH3, CO2, H2O), ок. 10% всей эн. – за счет а/к. (3) синт. N-соединений (аз. осн., нклтд, NAD, NADP, креатин, порфирины, ансерин, биогенные амины (КА, 5-НТ, Hyst). Азотистый баланс. а/к, б. – 90% N организма. Аз. бал. – разность между кол-вом поступившего и экскретированного N. Выд. N в осн. с мочой (15-16 г/сут). 2% выд. через кожу и фекалии. Аз. равновесие: поступ. = выд. → вес не изменяется. (+) Аз.б.: активн. рост, развитие, выздоровление, беременность. (–) Аз.б.: голодание, заб. с распадом тк. (СД, интексикация, гипертиреоз), старческий возраст. Если общ. азот мочи =100%, то мочевина =85%, креатинин =4-6%, мочевая к-та =1,6%, аммонийные соли =3%, неопределенная фракция =5%. Разрушение б.: убиквитиновая сис-ма. Убкв (8 кДа) отпр. повр. б. в протеасому (АТР-зав. активация). Общ. пути катаболизма а/к: (1) Переаминирование. Ферм. – аминотрансфераза, коферм. – пиридоксальфосфат (пр. вит.В6), мех-м пинг-понг. (2) Дезаминирование. 1) Восстановительное: R-CH(NH2)-COOH –(+2H)→ R-CH2-COOH + NH 2) Гидролитическое: R-CH(NH2)-COOH –(+Н2О)→ R-CH(ОН)-COOH + NH3 3) Внутримолекулярное: R-CH2-CH(NH2)-COOH –→ R-CH=CH-COOH + NH3 4) Окислительное: R-CH(NH2)-COOH –(+1/2 O2)→ R-C(О)-COOH + NH3 ! Окисл. дезамин. – осн. для чел-ка и жив. (исключительно с Glu с пом. глутамат ДГ): Glu + H2O ←(NAD(P)+ → NAD(P)H+H+)→ α-КГ + NH3. Подробнее: Glu (–2H) –(ГлДГ)→ HOOC-CH2-CH2-C(=NH)-COOH –(+H2O)→ HOOC-CH2-CH2-C(=O)-COOH + NH3 Регуляция: (+) ADP, GDP; (–) ATP, GTP. Обратно – восстановит. аминирование. (3) Декарбоксилирование. Судьба азота а/к: получ. в р-иях дезаминиования - рег. ферм. дезаминаза; декарбоксилазы → амины; переаминирование. Источники NH3: (1) неокисл. дезамин. нек. а/к (Сер, Тре, Гис) – в печени; (2) окисл. дезамин. Глу во всех тк. (кроме мыш. тк.); (3) дезамин. Глн и Асн – в печени и почках; (4) катаболизм биогенных аминов – во всех тк., в наиб. ст. в нервн. тк.; (5) ж/деят. МО толст. киш.; (6) распад пуриновых и пиримидиновых осн. Детокс. NH3: NH3 (все тк.) + Gln → Gln (р-ия амидирования, ферм. глутаминсинтетаза) NH3 (печень) + СО2 → цикл синт. мочевины → мочевина + фумарат; фумарат → ЦТК NH3 (почки) + Н+ → NH4+ (аммониогенез, ферм. глутаминаза.) Глн – трансп. форма аммиака в крови. Глн – осн. донор N для анаболитч. проц., идет на синт. нклтд, Glc, б., аминосахаров, Асн. Р-ия аммониогенеза обеспеч. сохр. важн. катионов. При ацидозе обр. NH4+ нейтрал. к-ты и в виде аммон. солей секрет. с мочой → защита от потери Na+ и K+ (при ацидозе ↑кол-во глутаминазы). Детокс. NH3: первичная, конечная. Судьба азота а/к:
(53) Детоксикация аммиака в организме. Цикл синтеза мочевины. Сумм. ур-ие: СО2 + NH3 + Asp + 3ATP + 2H2O → Мочевина + Фумарат + 2(ADP+Pi) + (AMP+PPi) карбамоилфосфат H2N-C(O)-OPO3H2 орнитин NH2-(CH2)3-CH(NH2)-COOH цитруллин (кето) NH2-C(O)-NH-(CH2)3-CH(NH2)-COOH цитруллин (енол) NH=C(OH)-NH-(CH2)3-CH(NH2)-COOH Asp NH2-CH(CH2-COOH)-COOH аргининосукцинат COOH-CH(NH2)-(CH2)3-NH-C(=NH)-NH-CH(COOH)-CH2-COOH фумарат COOH-CH=CH-COOH Arg NH2-CH[(CH2)3-NH-C(=NH)-NH2]-COOH мочевина NH2-C(O)-NH2 (1 ключ!) КарбамоилФосфатСинтетаза I NH3 + CO2 + 2ATP + H2O → карбамоилфосфат + 2ADP + Pi лок. в мтх гптц; N-ацетилглутамат – аллостерич. эффектор (без него акт-ть ферм. низкая), ↑сод. Glu в пище → стим. цикла синт. мочевины. (2) ОрнитинКарбамоилТрансфераза орнитин + карбамоилфосфат → цитруллин (кето) + Pi цитруллин (кето) → в ц/пл → цитруллин (енол) (3) АргининоСукцинатСинтетаза цитруллин (енол) + Asp + ATP → аргининосукцинат + AMP + PPi (4) АргининоСукцинатЛиаза аргининосукцинат → Arg + фумарат (5) Аргиназа Arg + H2O → мочевина + орнитин
В мол. мочевины атомы N: из мол. NH3, из Asp, – С – из СО2. NH3 обр. при окисл. дезаминир. Glu → NAD (затем→ 3АТР). Т.о., процесс синт. мочевины энергоНЕзатратный. ! Окисл. дезамин. – осн. для чел-ка и жив. (исключительно с Glu с пом. глутамат ДГ): Glu + H2 O ←( NAD( P)+ → NAD( P) H+ H+)→ α-КГ + NH3. Подробнее: Glu (–2 H) –(ГлДГ)→ HOOC- CH2- CH2- C(= NH)- COOH –(+ H2 O)→ HOOC- CH2- CH2- C(= O)- COOH + NH3 Регуляция: (+) ADP, GDP; (–) ATP, GTP. Обратно – восстановит. аминирование.
Фумарат → (в ЦТК) оксалоацетат → 1) дальше в ЦТК, 2) переаминир. в Asp (возм. вновь в синт. мочевины), 3) в Glc-неогенез.
(54) Болезни, вызванные генетическими дефектами ферментов цикла синтеза мочевины. Методы определения концентрации мочевины в крови и моче. Причины формирования и диагностическое значение азотемии. Дефекты цикла: при полн. отстуств. одного из ферм. – гибель в утробе матери. (1) КарбамоилФосфатСинтетаза I → тип I гипераммониемии NH3 + CO2 + 2ATP + H2O → карбамоилфосфат + 2ADP + Pi (2) ОрнитинКарбамоилТрансфераза → тип II гипераммониемии орнитин + карбамоилфосфат → цитруллин (кето) + Pi цитруллин (кето) → в ц/пл → цитруллин (енол) (3) АргининоСукцинатСинтетаза → цитруллинурия цитруллин (енол) + Asp + ATP → аргининосукцинат + AMP + PPi (4) АргининоСукцинатЛиаза → аргининосукцинатная ацидемия аргининосукцинат → Arg + фумарат (5) Аргиназа → гипераргининемия Arg + H2O → мочевина + орнитин Цитруллин и аргининосукцинат в N отсутствуют в крови! №1 и №2 – наиб. тяжелые. Причины токсичности аммиака – ?. Возм., нед. α-КГ. Либо угнет. р-ии переаминир. Сниж. ско. ЦТК. В крови – алкалоз → ↑сродство HHb к О2 → гипоксия. Либо пов. осмотич. давл. за счет высок. синт. Gln. → судороги, тремор, кома, смерть. Если б-нь распозн. быстро, то лечение простое: низкопротеиновая диета с доб. цитруллина и Arg. В кач-ве ЛС исп. бензоат Na (обр. компл. с Glu и Gly → NH3 пойдет на синтез). Предельно допустимый ур-нь аммиака в крови 60 мкмоль/л. Методы определения мочевины: (1) Ксантгидроловые: гетероцикл. спирт ксантгидрол → в р-ию с мочевиной → нераств. диксантилмочевина (опр. грави-, нефело-, колори- или титрометрически). Точно, но трудоемко. (2) Гипохлоритные – метод Б.А.Рашкована, появл. окраски при вз/д мочевины с гипохлоритом Na и фенолом, практич. не примен. (3) Диацетилмонооксимные: р-ия Фирона – вз/д мочевины и диацетилмонооксима с обр. окраш. прод., хорошая воспроизводимость, высокая чув-ть, большая специфичность. 2 этапа: 1. Гидролиз диацетилмонооксима с обр. диацетила и гидроксиламина. 2. Гидроксиламин вз/д с мочевиной с обр. окраш. диазинового пр. (4) Полуколичественные: с исп. индикаторной бумаги. (5) С исп. ионоселективных электродов. (6) Ферментативные: осн. на гидролизе мочевины уреазой (оптимум pH 6,0‑8,0) → аммиак опр. с пом. разл. р-ий (фенолгипохлоритная, с пом. глутаматДГ, салицилатно‑гипохлоритная). (7) Газометрические (гипобромитные) осн. на окисл. и разлож. мочевины гипобромитом Na в щел. среде: NH2-C(O)-NH2 + 3NaBrO → N2 + CO2 + 3NaBr + H2O. Выд. CO2 поглощает р-р, свободн. остается N2, объем которого измеряют. Метод неспецифичен (гипобромит реаг. с в-вами, сод. аминогруппы), неточен, плохо воспроизводим, трудоемок. Унифицир. методы: фенолгипохлоритный, диацетилмонооксимный и уреазный методы, экспресс‑метод с использованием индикаторной бумаги "Уреатест". N величины: Сыворотка (по р-ии с диац.моноокс.; с реакт. Несслера): взр. 2,5–8,3 ммоль/л, (по р-ии с фенолгипохлоритом) 3,3–8,3 ммоль/л. Моча (по р-ии с диац.моноок.) 330–580 ммоль/сут Моча (по р-ии с реакт. Несслера) 430–710 ммоль/сут Слюна ок. 1,83 ммоль Для сопоставл. концентр. остат. азота с сод. азота мочевины концентр. последней разделить на 2,14. Сыворотка. Ур-нь мочевины зав. от ск. ее синт. в печени и выд. через почки, а также от величины б. катаб. ↑ уровня: при нар. ф-ии почек, истощ. запасов воды, повыш. катаб. б. (о. инф. миок., стресс, ожоги, желт. атроф. печени, ЖК кр/теч), при диете с выс. сод. б. ↓уровня: при диете с низк. сод. б., при тяж. заб. печени, сопр. нар. синт. мочевины (паренхим. желтуха, цирроз). Моча.↑кол-ва: гипертиреоз, злокач. анемия, лихорадка, при отрав. фосфором, при диете с выс. сод. б., в послеоперац. п-оде. ↓кол-ва: нефрит и др. заб. почек, паренхим. желтуха, цирроз или дистроф. печени, у здоровых растущих детей и при низкобелк. диете.
(55) Общие реакции обмена аминокислот. Кетогенные и глюкогенные аминокислоты. Общ. пути катаболизма а/к: (1) Переаминирование. Ферм. – аминотрансфераза, коферм. – пиридоксальфосфат (пр. вит.В6), мех-м пинг-понг. (2) Дезаминирование. 1) Восстановительное: R-CH(NH2)-COOH –(+2H)→ R-CH2-COOH + NH 2) Гидролитическое: R-CH(NH2)-COOH –(+Н2О)→ R-CH(ОН)-COOH + NH3 3) Внутримолекулярное: R-CH2-CH(NH2)-COOH –→ R-CH=CH-COOH + NH3 4) Окислительное: R-CH(NH2)-COOH –(+1/2 O2)→ R-C(О)-COOH + NH3 ! Окисл. дезамин. – осн. для чел-ка и жив. (исключительно с Glu с пом. глутамат ДГ): Glu + H2O ←(NAD(P)+ → NAD(P)H+H+)→ α-КГ + NH3. Подробнее: Glu (–2H) –(ГлДГ)→ HOOC-CH2-CH2-C(=NH)-COOH –(+H2O)→ HOOC-CH2-CH2-C(=O)-COOH + NH3 Регуляция: (+) ADP, GDP; (–) ATP, GTP. Обратно – восстановит. аминирование. (3) Декарбоксилирование. Судьба азота а/к: получ. в р-иях дезаминиования - рег. ферм. дезаминаза; декарбоксилазы → амины; переаминирование. Детокс. NH3: (1) NH3 (все тк.) + Gln → Gln (р-ия амидирования, ферм. глутаминсинтетаза) (2) NH3 (печень) + СО2 → цикл синт. мочевины → мочевина + фумарат; фумарат → ЦТК (3) NH3 (почки) + Н+ → NH4+ (аммониогенез, ферм. глутаминаза).
Декарбоксилирование. NH2-CH(R)-COOH –(субстрат-декарбоксилаза)→ R-CH2-NH2 + CO2 Декарбоксилазы раб. с коферм. пиридоксальфосфатом (вит.В6). Амины – ядов. в-ва, должны выводиться из организма (орнитин → путресцин, Lys → кадаверин, т.е. трупные яды); явл. субстр. для синт. разл. в-в; биол. акт. амины. Катаболизм углер. скелета а/к: кето/глюкогенные. (1) кетогенные. Leu, Lys – чисто кетогенные. Leu, Trp, Ile → AcCoA → AcAcCoA→ кет. тела; Leu, Lys, Trp, Tyr, Phe → AcAcCoa → кет. тела. (2) глюкогенные. Gly, Ala, Val, Pro, Ser, Tre, Cys, Met, Asp, Asn, Glu, Gln, Arg, His – чисто глюкогенные. Ala, Ser, Gly, Cys, Tre, Trp → ПВК → Glc; Asn → Asp → ЩУК → Glc; Tyr, Phe, Asp → Фумарат → ЩУК → Glc; Val, Ile, Met → Сукцинил-КоА → Фумарат → ЩУК → Glc; Arg, Pro, His, Gln, Glu → α-КГ → Сукцинил-КоА → Фумарат → ЩУК → Glc. ЩУК, Фумарат, Сукцинил-КоА, α-КГ – интермедиаты ЦТК. (3) смешанные. Ile, Phe, Tyr, Trp.
(56) Реакция декарбоксилирования аминокислот. Синтез биогенных аминов: гистамина, серотонина, катехоламинов. Общий путь распада биогенных аминов. Декарбоксилирование. NH2-CH(R)-COOH –(субстрат-декарбоксилаза)→ R-CH2-NH2 + CO2 Декарбоксилазы раб. с коферм. пиридоксальфосфатом (вит.В6). Амины – ядов. в-ва, должны выводиться из организма (орнитин → путресцин, Lys → кадаверин, т.е. трупные яды); явл. субстр. для синт. разл. в-в; биол. акт. амины. Для всех аминов раб. ферменты МАО и ДАО: R-CH2-NH2 + H2O + O2 → R-CHO + NH2 + H2O2 APUD-system (Amine Precursors Uptake and Decarboxilation): ок. 60 типов кл., мн. в киш. (энтерохромафф. кл.). Поглощ. а/к-предшеств. → амины. Амины≠гормоны (действие короткое). Гистамин. Синт. из His, ферм. ГистидинДеарбоксилаза. Больше всего в коже, легких, печени, ЖКТ. Депо в mast cells и БФ. Ф-ии: медиатор ЦНС, ваходилат., бронхоконстр., восп., стим. секр. HCl. Живет недолго, разрушение в печени. Ферменты гистаминаза и альдегидоксидаза: (1) Гистамин → метилгистамин → 1-метил-4-имидазол-ацетальдегид → метилимидазолуксусная к-та (2) Гистамин → имидазолацетальдегид → ИУК (имид.укс. к-та)
Серотонин(5-гидрокситриптамин, индолалкиламин). Синтез: Trp –(Trp-гидроксилаза)→ 5-ОксиТриптофан –(декарбоксилаза)→ 5-НТ. Ф-ии: нейромед., мощн. вазоконстр., сокр. ГМК, боль, восп., облад. седат. действ. (1) 5-НТ –(N-ацетилаза)→ N-ацетилсеротонин –(N-ацетилсеротонин-О-метилтрансфераза)→ мелатонин. Впервые выд. из эпифиза, синт везде, выраб. только ночью, обл. антигонадотропн. действ. (2) 5-НТ –(МАО)→ 5-ОксиИндолУксусный альдегид –(альдегидгидроксилаза)→ 5-ОксиИндолУксусная к-та. (3) 5-НТ –(N-метитрансфераза)→ N-метилсеротонин –(N-метилтрансфераза)→ буфотенин. ↑АД, стим. выд. Adr, актив. психоэмоц. сферу.
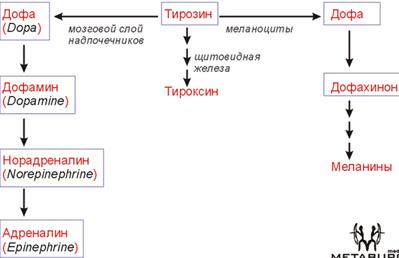
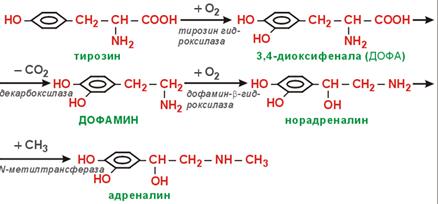
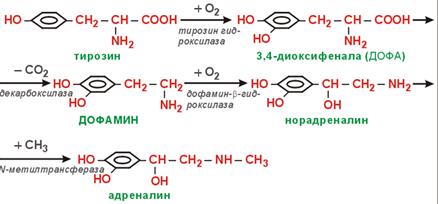
КА. Tyr (в мозг. слой н/поч) → Dopa → Dopamine → Nadr → Adr. Катаболизм КА: 1) исп. МАО (классич. путь); 2) индивид. для Adr.
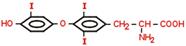
(57) Обмен фенилаланина и тирозина. Болезни, вызванные генетическими дефектами ферментов обмена этих аминокислот. Phe → на синт. б. (незам. а/к); → Tyr. Phe –(ФенилаланинГидроксилаза)→ тирозин –(ТирозинАминоТрансфераза)→ 4-ГидроксиФенилПируват –(оксидаза 4-ГФПВК)→ гомогентизиновая к-та –(оксидаза гомогентизата)→ 4-МалеилАцетоАцетат –(МалеилАцАц Изомераза)→ 4-ФумарилАцАц –(Гидролаза ФумарилАцАц)→ Фумарат + АцАц ФенилаланинГидроксилаза. +О2; Н4-биоптерин. Х→Фенилкетонурия. Phe токсичен, только диетич. коррекция. ТирозинАминоТрансфераза. α-КГ→Glu. Х→Тирозинемия, тип 2. оксидаза 4- ГидроксиФенилПирувата. Вит.С; гемовое Fe. Х→Неонатальная тирозинемия. Измен. кожн. покр., нар. зрения. оксидаза гомогентизата. Х→Алкаптонурия. Гомогентензиновая к-та отклад. в СТк, хрящ. тк. В моче превращ. в алкаптон (моча темная, коричн.). МалеилАцАц Изомераза. Гидролаза ФумарилАцАц. +Н2О (в гптц). Х→Тирозинемия, тип 1. Лет., т.к. накапл. 4-ФумарилАцАц (12-15 лет). Накопление Tyr не опасно, т.к. Tyr имеет разл. пути метаб. (в отл. от Phe): → Т3, Т4; →КА; →меланины; →фумарат; →АцАц.
Tyr: см. 58.
(58) Биологически активные производные тирозина. Локализация синтеза и их роль в организме.
КА. Tyr (в мозг. слой н/поч) → Dopa → Dopamine → Nadr → Adr. Катаболизм КА: 1) исп. МАО (классич. путь); 2) индивид. для Adr.
Катаболизм:
Меланины. Биосинт. в меланоцитах. Tyr –(тирозиназа(Cu2+, Альбинизм – дефект по тирозиназе. Меланины: эу-/фео-.
Т3 и Т4. Предш. – тироглобулин (115 остатков Tyr; 660 кДа; накапл. в коллоиде). Этапы синт.: 1) конденсация йода при уч. Na+/ K+- ATPase. 2) окисление йода (только в щит. жел., ферм. тиреопероксидаза): J(–) + NADPH+H+ + H2O2 –(тиреопероксидаза)→ I(+) +NADP+ 2Н2О. 3) органификация йода. 4) →Т3, Т4. +для Т4→Т3 Se-зав дейодиназа.
(59) Биосинтез креатина, креатинфосфата в организме. Диагностическое значение креатина и креатинина. Карнитин, карнозин, ансерин. Их роль в организме. Креатин. 1я стадия – в почках, 2я – в гптц. (1) Arg + Gly –(трансамидиназа)→ гуанидинацетат + орнитин NH2-C(=NH)-NH-(CH2)3-CH(-NH2)-COOH + NH2-CH-COOH → NH2-C(=NH)-NH-CH2-COOH + NH2-(CH2)3-CH(-NH2)-COOH (2) Гуанидининацетат + -СН3 –(S-аденозилметионин→S-аденозилгомоцистеин)→ креатин (МетилГуанидинУксусная к-та) NH2-C(=NH)-NH-CH2-COOH + -CH3 → NH2–CH(=NH)–N(-CH3)–CH2–COOH NH2–CH(=NH)–N(-CH3)–CH2–COOH + ATP ←(креатинкиназа)→ H2O2P~NH–CH(=NH)–N(-CH3)–CH2–COOH + ADP Креатинин:
Креатин → в кровь, далее захв. кл. мыш. и нервн. тк., где АТР-зав. → в креатинфосфат → быстро мобилизуемая энергия (на 5-10 сек). Показатель почечной фильтрации – клиренс креатинина. Трансамидиназа – индикаторный ферм. почечной тк. (в N в моче нет). Креатинкиназы: КК-МВ (сердечный), КК-ВВ (мозговой), КК-ММ (СкМ). Карнитин. Синт. интенс. в печени, почках. Нужны: Lys, аденозил-Met, аскорбиновая к-та, Fe. Карнитин – транспортер ЖК из ц/пл в мтх. БАВ, витаминопод. фактор (часто бывает мало). (CH3)3-N(+)-CH2-CH(OH)-CH2-COO(–) Карнозин (β-аланилгистидин), Ансерин (β-аланил-метилгистидин), Гомокарнозин (γ-аминобутирилгистидин) – антиокс., увелич. буф. емкость мышц при сокр., уд. ионный баланс. Гомокарнозина мало, карнозина много.
(60) Синтез и катаболизм гема. Значение конъюгирования продуктов метаболизма гема в печени.
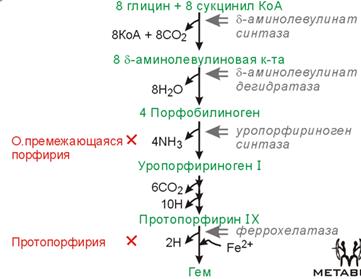
Хромопротеины: флавопроетины, гемопротеины (Hb, миоглобин, каталаза, пероксидаза, цитохромы). Гем синт. во всех кл. Гем – протопорфириновое кольцо с разными R и ионом внутри. Сопряженная, оч. уст. стр-ра. Пиррольные кольца соединены метеновыми мостиками. Fe имеет 2 ковал. св. с N, 2 координационные св. (дон.-акц. мех-м) с N, 3я коорд. св. – с мол. О2 (перпендик.), 4я коорд. св. – с N гистидина глобина (перпендик.). Синтез: Ключ. ферм. – δ-аминолевулинатсинтаза → регул.: ингиб. прямой обр. св. гемом (Fe2+), гемином (Fe3+). Pb – мощн. ингибитор, As, Hg – менее. Барбитураты, сульфаниламиды – то же.
ПОРФИРИЯ – общ. назв. для заб., связ. с наруш. синт. гема. Клин. симпт.: в осн. неврология, абдоминальные боли, выд. большого кол-ва субстратов (моча темнеет на воздухе). Наиб. тяж. – ПРОТОПОРФИРИЯ. Симпт.: крапивница, пов. чувств. к свету. Стр-ра ПОРФИРИНОВ:
О-гем – не удалены до конца Н, связь между пирролами – метиленовые мостики (не имеют сопр. связей, не явл. хромогенами). КАТАБОЛИЗМ гема: живет до 120 дн. 2*10^11 ЭрЦ/сут разруш. Если ЭрЦ разруш. в русле крови, то Hb будет связан гаптоглобином (в составе гаптогл. всегда метгемоглобин, т.к. Fe2+ оч. быстро Ох). Если протеиназы крови работают до связ. с гаптоглоб., то высвоб. гем, кот. улавливается гемопиксином. Если своб. Fe оказ. в кр. русле, то его потере препятств. трансферрин. Fe связ. в печени ферритином (оч. большой, 24 с/ед, может св. ок. 2500 мол.). Билирубин: 87% - распад гема, 13% - распад др. гемопротеинов. Билируб. обр. в печени, выводится с желчью в 12пК.
Билирубин сам выд. не может, осн. часть обр. в кл. РЭС селезенки и ККМ → в кровь (перенос альбуминами). БНА (Билирубин Неконъюгированный, соединенный с Альбуминами) = свободный = непрямой. УДФ-глюкуроновая + Билирубин –(УДФ-глюкуронилтрансфераза)→ билирубин-глюкуронид + УДФ БК (обр. только в печени) = БМГ + БДГ. При гепатитах БМГ больше. БК хорошо раств., легко выводится желчью, легко → в кровь. N 0,1–4 мкМ/л. ОБ = БНА+БК. N до 20 мкМ/л. БНА/БК – осн. диагностич. показ. В желчи мн. БК, мало БНА. Если плохо раб. трансфераза, то больше БНА, обр. билирубинат Ca → ↓в виде камней.
(61) Причины гипербилирубинемии, виды желтух. Диагностическое значение общего, прямого и непрямого билирубина. Метод определения билирубина в крови. В желч. пуз. из БК → отщ. глюкуроновая к-та → Red до уробилиногена (в желч. пуз., тонк. киш.), стеркобилиногена (в ниж. отд. киш.). → Уро-/Стеркобилины= УРОБИЛИНОИДЫ (выв. с фекальными массами, 40-280 мг/сут). Часть уробилиноидов всас.: уробилиноген (в тонк. к.) → воротная вена → печень → разруш. до пирролов → выв. с желчью (ЭнтероГепатич. уробилиногеновый цикл). Стеркобилиноген → геморроидальные вены → общ. кровоток → почки. О,6-4,0 мг/сут – уробилиноиды мочи. БИЛИРУБИНУРИЯ св. с БК. Причины: восп. в печени (N БК в крови мало). ЖЕЛТУХИ.
ДЛЯ ИССЛ ОБ: 1. Прямые спектрофотометрические методы, осн. на измер. абсорбции билирубина при 440‑460 нм, источником ошибок здесь является интерференция желтых небилирубиновых пигментов; 2. Ферментативные методы, осн. на окислении пигмента медьсодержащим ферментом билирубиноксидазой (КФ 1.3.3.5.), с образованием биливердина и исчезновением абсорбции при 460 нм. Методы этой группы точны и отличаются высокой специфичностью и чувствительностью; 3. Колориметрические диазометоды, базируются на взаимодействии билирубина с диазотированной сульфаниловой кислотой с образованием азопигментов. Под воздействием кислоты разрывается тетрапирроловая структура билирубина с образованием двух дипирролов, углеродные атомы метиленовых групп вступают в прямую реакцию с диазотированной сульфоновой кислотой (диазосмесь) с образованием розово-фиолетовых изомеров азодипиррола с максимумом поглощения при 530 нм. Связанный билирубин реагирует быстро, несвязанный –– только после добавления акселератора (кофеин, метанол, мочевина, бензоат или гидроокись натрия, уксусная кислота и другие). Последний освобождает билирубин из комплекса с белками и тем самым ускоряет реакцию азосочетания. Образовавшийся азокраситель ведет себя как кислотно-основной индикатор с несколькими цветными переходами: в сильнокислой среде он окрашен в фиолетовый цвет, в слабощелочной и слабокислой — в розовый, в сильнощелочной среде — в синий или зеленый цвета. 4. Электрохимические методы с использованием платинового и ртутного электродов; 5. Хроматографическое разделение отдельных фракций билирубина; 6. Флюорометрические методы, используется свойство свободного билирубина после поглощения при 430 нм испускать свет длиной 520 нм. Позволяют определить концентрацию неконъюгированного билирубина и, после обработки детергентом, общего билирубина главным преимуществом метода является возможность использовать микроколичества сыворотки. Унифицированным методом является метод Йендрашика‑Клеггорна‑Грофа по диазореакции в щелочной или слабокислой среде в присутствии акселератора.
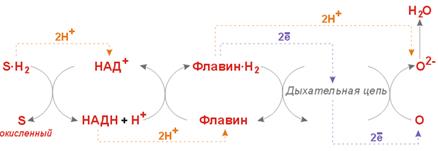
6. ЭНЕРГЕТИЧЕСКИЙ ОБМЕН 62. Общие и специфические пути катаболизма белков, жиров и углеводов (по Кребсу). Виды биологического окисления. Человек и животные – гетеротрофы. Метаболизм: аэробный, анаэробный. 1 моль Glc (180 г) → анаэр:45’000 кал, аэр:686’000 кал. Энергетич. обмен – трансформация Е хим. в-в в Е АТР или др. конвертир. формы и исп. кл. этих форм Е для соверш. работы. Конвертир. формы Е: АТР, ΔμН+, Δμ Na +. Пища – источник Е, но она почти не сод. первичных доноров Н+ (в-в, кот. легко м.б. окислены) и восстановленных эквивалентов. Перв. доноры: ПВК, АсСоА, α -КГ, сукцинил-СоА, фумарат, оксалоацетат. Стадии катаболизма: Биологическое Ох – Ох орг. в-в в живом организме; –сов-ть б/х р-ий, происх. в жив. орг., в кот. орг. в-ва через ряд посл. р-ий подв. Ох; – сложный долгий многоступенч. процесс, в кот. участв. Red - Ox ферменты (в он., ДГ). ДГ: никотиновые ( NADHH + более энергоемкое соед, чем FADH 2), флавиновые ( FAD – более сильный восстанов., чем NAD +). Синтез АТР : ADP + Pi –(+ E )→ ATP + H 2 O В орг. Ох, как правило, сопряжено с фосфорилированием → окислительное фосфорилир. (идет на внутр. мембр. мтх; синт. АТР из ADP и Pi , сопряж. с Ох субстр. и переносом е по дых. цепи). (23) Окислительное декарбоксилирование пирувата. Пируватдегидрогеназный ферментный комплекс. ПВК + CoA-SH + NAD+ –(PDH)→ AcCoA + CO2 + NADH+H+ |
Последнее изменение этой страницы: 2019-03-31; Просмотров: 187; Нарушение авторского права страницы