|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Эпигенетические модификации. Модификации ДНК.
Эпигенетика – наука о наследуемых свойствах организма, которые не связаны с изменением собственно нуклеотидной последовательности ДНК и могут быть не прямо, а опосредованно закодированы в геноме. К числу известных эпигенетических механизмов (сигналов) относятся: энзиматическое метилирование ДНК, гистоновый код (разные энзиматические модификации гистонов – ацетилирование, метилирование, фосфорилирование, убиквитинирование и др.) и замалчивание генов малыми РНК (miRNA, siRNA).» ([11]). Метилирование ДНК Теперь что представляет собой метилирование ДНК. Это результат процессов, в ходе которых на определённые нуклеотиды в составе ДНК садится или же наоборот удаляется метильная группа. Такая метильная группа может исполнять роль метки-закладки, информирующей белковые комплексы как им себя вести относительно этого участка ДНК. Например, если метильные метки расположены на промоторе (последовательности нуклеотидов ДНК на которую садится РНК-полимераза чтобы начать синтез РНК) гена, то это обычно приводит к ингибированию (репрессии) данного гена. Удаление с промотора метильных групп (деметилирование) приводит к активации (дерепрессии) такого гена. ДНК построена из четырёх нуклеотидов, но у эукариот метилированию обычно подвергается только один из них – цитозин. Посадку метильной группы осуществляет фермент ДНК-метилтрансфераза. Следует при этом отметить, что регуляция посредством метилирования ДНК характерна не для всех организмов. Так, например у плодовой мушки дрозофилы число таких метилированных цитозинов ничтожно мало – всего 0.03%, а у червячка C. elegans и дрожжей и того меньше ([21]). У мышки их заметно больше – порядка 4%. Казалось бы так ли уж важно понимать процессы метилирования ДНК для того чтобы разобраться в механизмах старения, ведь у плодовой мушки и у дрожжей эти процессы не играют существенной роли, однако они стареют также как и другие организмы. Почему важно изучать роль метилирования ДНК в механизмах старения? Начнём с того, что метилирование ДНК как правило, исчезает при образовании зиготы – клетки, образующейся в результате оплодотворения и с которой начинается процесс развития нового организма. Затем, по мере развития организма, происходит постепенное накопление всё новых и новых метильных групп. Причем характер метилирования, регулирующего генную активность (экспрессию) при делении клеток передается вновь образованным клеткам. Считается, что метилирование ДНК позволяет стабилизировать экспрессию генов в клетках, закрепляя «клеточную память» в клетках, которые делятся и дифференцируются из эмбриональных стволовых клеток в клетки различных специализированных тканей организма. Поскольку изменение метилирования ДНК закреплено клеточной памятью и стабильно наследуется клетками-потомками данной клетки, но не является само по себе мутацией ДНК, его можно назвать эпигенетическим. ([22]) Все соматические клетки организма (за исключением иммунных клеток вроде T и B клеток, а также эритроцитов, которые при созревании вообще теряют ядро), несмотря на значительные различия в их физиологических функциях, имеют ДНК с одной и той же последовательностью нуклеотидов, но отличающуюся по характеру метилирования. Этот феномен — что в разных клетках одного и того же организма ДНК метилирована по-разному, Ванюшин с соавторами обнаружили ещё в 1970 м году, что уже тогда позволило им первыми заявить, что метилирование ДНК – механизм регуляции экспрессии генов и клеточной дифференцировки ([23]). Было обнаружено что после рождения в ходе старения у коровы, крысы и у мышей понижается содержание метилированного цитозина, причем понижение это происходит главным образом на участках ДНК, несущих многократно повторяющиеся последовательности ([24] [25]). При этом была подмечена закономерность — Mus musculus теряет участки метилирования со скоростью порядка 4,7 Х 104 (в месяц примерно 0.012% от уровня новорожденного), тогда как Peromyscus leucopus теряет 5-метилцитидин со скоростью 2.3 Х 104 (в месяц примерно 0.006% от уровня новорожденного) и при этом живет в два раза дольше чем Mus musculus. У человека общий уровень метилирования ДНК в соматических клетках также неуклонно снижается. В фибробластах человека при приближении репликативного старения, когда способность клетки к делениям заметно падает, происходит общее снижение уровня метилирования ДНК (главным образом в районах бедных на содержание генов, поздно реплицирующихся участков, а также LAD (lamina-associated domain) – доменах ДНК примыкающих к ядерной мембране (что сопровождает отслаивание гетерохроматина от ядерной ламины и замену ламина А на его укороченный аналог – прогерин ([26]) ), Все эти участки обычно несут метки гистонов неактивного хроматина — гетерохроматина). Вместе с тем происходит очаговое гиперметилирование там, где расположены CpGi островки, локализованные на генных промоторах, где метилирование ДНК приводит к ингибированию генов. По мнению авторов этого исследования и их последователей, постаревшие клетки очень похожи на раковый эпигеном ([27] [28]).
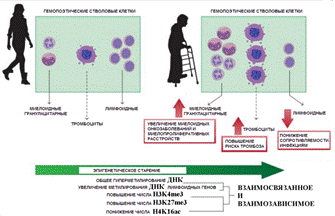
По статье ( ) Эпигенетические изменения при старении гемопоэтических стволовых клеток, приводят к увеличению числа случаев возрастных миелопролиферативных заболеваний и злокачественных опухолей миелоидного типа, а также к повышенному риску тромбоза сосудов и снижению сопротивляемости организма к инфекции. В отличие от других клеток организма метилирование ДНК при старении гемопоэтических стволовых клеток не понижается, а наоборот повышается Однако в стволовых клетках предшественниках клеток крови, которые сохраняют способность к делению до глубокой старости организма метилирование ДНК не снижается, а перераспределяется: усиливается в участках генома связанных с лимфоидной и эритроидной специализацией. В результате при старении количество образуемых клеток этих типов снижается ([29]). |
Последнее изменение этой страницы: 2019-04-19; Просмотров: 234; Нарушение авторского права страницы