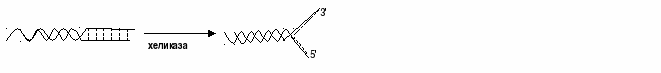|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Ядро. Строение и свойства.
Ядро клетки — главный центр с генетической информацией, так как в нем находятся хромосомы, содержащие наследственные признаки, закодированные в форме ДНК. Другие носители информации имеют меньшее значение. Положение, форма и размеры ядра могут изменяться, часто параллельно с изменениями интенсивности метаболизма. Ядро чаще всего расположено в центре клетки, и только у растительных клеток с центральной вакуолью — в пристеночной протоплазме. Оно может быть различной формы: · сферическим; · яйцевидным; · чечевицеобразным; · сегментированным (редко); · вытянутым в длину; · веретеновидным, а также иной формы. Диаметр ядра варьирует в пределах от 0,5 мкм (у грибов) до 500 мкм (в некоторых яйцеклетках), в большинстве случаев он меньше 5 мкм. Ядро состоит из: · нуклеоплазмы; · хромосом (хроматина); · ядрышек; · ядерной оболочки, представляющей собой часть эндоплазматического ретикулума. Клеточные ядра образуются только из ядер. Репликация ДНК, т. е. удвоение генетической информации, гарантирует идентичность ядер, несмотря на всю сложность их деления. Главные функции клеточного ядра следующие: · хранение информации; · передача информации в цитоплазму с помощью транскрипции, т. е. синтеза переносящей информацию и-РНК; · передача информации дочерним клеткам при репликации — делении клеток и ядер.
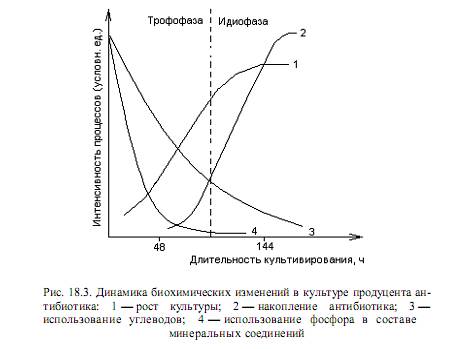
Биосинтез антибиотиков Антибиотики относятся к вторичным метаболитам — низкомолекулярным веществам, функции которых несущественны для роста чистых культур. Вторичные метаболиты (антибиотики, алкалоиды, гормоны роста растений, токсины) производятся представителями ограниченного числа таксономических групп. В отличие от первичных метаболитов, вторичные имеют более сложное химическое строение и образуются, как правило, в ходе длинных метаболических путей. Известно, что образование антибиотиков делает их продуцентов более конкурентоспособными и является мощным фактором в борьбе организмов за существование. Однако это не единственная роль антибиотиков: недавно обнаружено, что данные вещества осуществляют функцию контроля и регуляции процессов метаболизма, выполняя роль специфических эффекторов, контролирующих активность ряда ферментных систем. Кроме этого, существует точка зрения, согласно которой антибиотики могут служить клеточными компонентами, в виде которых клетка избавляется от накопившихся в ней по тем или иным причинам в избытке метаболитов и восстановительных эквивалентов. Биоосинтез большинства вторичных метаболитов, в том числе антибиотиков, обычно осуществляется на определенной стадии развития микроорганизма, в условиях торможения основных процессов анаболизма—биосинтеза нуклеиновых кислот, белка, липидов, полисахаридов. Различают две принципиально различные стадии развития популяции продуцентов антибиотиков. Первая, носящая название трофофаза или фаза сбалансированного роста, характеризуется интенсивным синтезом соединений, требующихся для роста клеток, что сопровождается быстрым потреблением основных питательных компонентов среды. Эта фаза соответствует фазе логарифмического роста периодической культуры микроорганизмов. После исчерпания некоторых ингредиентов среды (в первую очередь, углерода, азота, молекулярного кислорода) и накопления продуктов метаболизма наступает вторая фаза (идиофаза, или фаза несбалансированного роста). Идиофаза отличается заметным торможением или прекращением роста микробной популяции, что соответствует переходу в стационарную фазу роста периодической культуры. Здесь начинают преобладать протеолитические процессы, происходит автолитический распад клеток. Именно в идиофазе многие микроорганизмы начинают синтезировать вторичные метаболиты, к числу которых относятся и антибиотики. Описанные закономерности можно отобразить графически (рис. 18.3).
В большинстве случаев максимум накопления антибиотика в культуральной жидкости наступает после максимума накопления биомассы. Это объясняется тем, что синтез ферментов, ответственных за образование антибиотиков, подавлен (репрессирован) во время трофофазы. Биосинтез антибиотиков — наследственно закрепленное свойство организмов. Гены, ответственные за эти процессы, могут располагаться на хромосомах (нуклеоидах у прокариот), а также на плазмидах. Регуляция процесса синтеза антибиотика осуществляется разными способами: чаще в ходе катаболитной репрессии, с помощью репрессии синтеза ферментов конечным продуктом, а также при аллостерическом ингибировании антибиотиком активности ключевых ферментов своего биосинтеза. При этом важными компонентами среды, оказывающими влияние на биосинтез антибиотиков, являются глюкоза, а также фосфат. Как правило, избыток этих веществ отрицательно сказывается на образовании антибиотиков. Антибиотическая продуктивность организма отражает количество антибиотика (в мкг или в единицах), образованное единицей массы (обычно 1 мг) сухих клеток микроорганизма-продуцента в единицу времени (обычно за 1 ч). Этот параметр зависит, в первую очередь, от вида микроорганизма, фазы роста клеток, состава среды, условий культивирования. Препараты антибиотиков характеризуются разной степенью чистоты и, соответственно, активности, которая выражается в количестве единиц антибиотической активности (ЕД), содержащихся в 1 мг или 1 мл препарата. За единицу антибиотической активности принимают минимальное количество чистого вещества антибиотика, способное подавить развитие определенного числа клеток стандартного штамма микроорганизма (тест-микроба) в единице объема культуральной жидкости. Например, 1 ЕД стрептомицина эквивалентна 1 мкг чистого основания этого вещества, 1 ЕД пенициллина соответствует 0,6 мкг натриевой соли бензилпенициллина. При этом в качестве тест-микроба для стрептомицина используется E.coli, а для пенициллина — Staphylococcus aureus. Синтез биополимеров Схематически все процессы образования полипептидов и полинуклеотидов укладываются в следующую схему:
- где над стрелками обозначены названия соответствующих стадий. Общие принципы реакций Все биополимеры (полипептиды, гетерополисахариды, полинуклеотиды) имеют организацию, - специфическую упаковку в пространстве, поэтому их синтез включает дополнительно и стадию структурирования. А если их звенья еще и отличаются друг от друга, отсюда, чтобы вновь образующийся полимер включал их в строго определенном порядке, необходима матрица, что и отличает генез белков и нуклеиновых кислот от других подобных процессов. Кроме того, любой синтез требует затрат энергии, причем часто в этих случаях используются не обычные макроэрги, а происходит с их помощью предварительная активация субстратов. Исходя из этих предпосылок, можно выделить следующие общие принципы, характерные для этих процессов: 1. Наличие матрицы: в синтезе дочерней ДНК ею служит вся материнская ДНК; для образования РНК используется фрагмент ДНК (ген, или транскриптон); аминокислотная последовательность определяется набором триплетов иРНК. 2. Субстраты предварительно активируются: на первой стадии используются дезоксирибонуклеотидтрифосфаты (дАТФ, дГТФ, дЦТФ, ТТФ), генез РНК требует наличия соответственно АТФ, ГТФ, ЦТФ, УТФ, полипептиды получаются из аминоацил-тРНК. 3. Основные ферменты, обеспечивающие собственно полимеризацию, относятся к классу трансфераз и называются ДНК-, РНК-полимеразы и пептидилтрансфераза. 4. Сложность образования биополимера выделять в этом процессе две фазы: собственно синтез (как бы начерно) и созревание – укладку в пространстве, химическую модификацию. 5. В собственно синтезе выделяют следующие стадии: а) инициацию – начало; б) элонгацию (longus- длинный) – наращивание цепи; в) терминацию – остановку реакций. Репликация ДНК Репликация – синтез дочерней ДНК с использованием в качестве матрицы всей молекулы материнской ДНК. С этого процесса и начинается деление клетки в S-фазу (синтетическую). В основе репликации лежат три принципа: а)комплементарности– азотистые основания образующейся дочерней цепи должны обладать химическим и геометрическим соответствием с подобными веществами материнской нити;полуконсервативности– каждая из синтезированных молекул ДНК состоит из одной материнской и одной дочерней; в)однонаправленности – считывание матрицы идет от 5' к 3' концу. Чтобы порядок азотистых оснований, находящихся внутри нитей ДНК, можно было считывать, необходимо разорвать водородные связи, соединяющие между собой цепи и лишить их спирализации. Для этих целей используются следующие ферменты: топоизомеразаихеликаза. Первая гидролизует на одном из участков полинуклеотида фосфодиэфирная связь, что позволяет этому фрагменту раскрутиться относительно параллельно л а затем восстановить связь. Хеликазаразрушает водородные взаимодействия между противолежащими основаниями участков двух цепей:
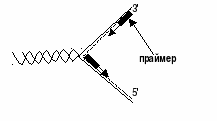
Под ее действием фрагменты расходятся, образуя репликативную вилку. Затем с помощьюпраймазы(primer- затравка) синтезируется олигорибонуклеотид – небольшое соединение, состоящее (обратите внимание) из 8-10 монорибонуклеотидов. Оно служит местом, куда крепится основной фермент –ДНК-полимераза. Из-за антипараллельности цепей данные структуры будут локализоваться в разных местах: одна у конца, другая несколько в глубине противоположной нити:
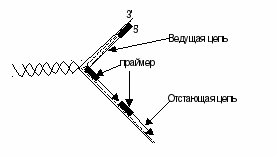
Все дело в том, что ДНК-полимеразыне могут начинать синтез, а способны только добавлять дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся цепи, чем и являетсяпраймер. Описано 3 класса данных энзимов: в ядрах присутствуют полимеразы альфа (Роlα), ответственные за хромосомную репликацию, полимеразы бета (Роlβ), которые используются при необходимости при репарации. Третий их представитель – ДНК-полимераза гамма (Роlγ) осуществляет синтез кольцевого генома митохондрий. Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом макроэргической связи в дНТФ и отщеплением пирофосфата. Важная деталь – генетический материал живых организмов имеет огромные размеры, но реплицируется с высокой точностью. В среднем при воспроизведении генома (ДНК длиной более 3 млрд пар нуклеотидов) возникает не более 3-х ошибок благодаря наличию специальных механизмов, осуществляющих необходимую коррекцию. Суть последней в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй – перед присоединением нового нуклеотида. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний мономер удлиняющейся дочерней нити ДНК образовал правильную (комплементарную) пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии произошло ошибочное спаривание оснований, то фермент возвращает последнее добавленное звено, после чего освободившееся место занимает правильный нуклеотид. Параллельно с ростом цепи продолжается деспирализация и разделение цепей с помощью топоизомеразы и хеликазы, удлинение репликативной вилки. А так как процесс осуществляется лишь в одном (5' – 3') направлении, что позволяет происходить этому явлению непрерывно только на одной из матричных нитей. На антипараллельной синтез ограничивается сравнительно короткими фрагментами (100-1000 мононуклеотидов), названными по имени обнаружившего их ученого фрагментами Оказаки(с постоянным предварительным синтезом праймеров):
Так продолжается до тех пор, пока вся матрица не выполнит свою функцию, причем во время репликации специальные нуклеазы вычленяют праймеры. Если позволяет направление, ДНК-полимеразы застраивают возникшие бреши, а сшивание сближенных фрагментов обеспечивается ДНК-лигазами– так завершается собственно синтез (стадия терминации). В тех концах дочерних цепей, где находились праймеры, которые позднее были удалены, не происходит достраивания дезоксирибонуклеотидами (невозможно считывание в обратном направлении), отсюда при каждом делении клетки молекулы новых цепей укорачиваются на 10-20 нуклеотидов, но объем информации при этом не уменьшается, так как потерянные участки ее не несли. Дело в том, что когда в эмбриональных тканях шли усиленные митозы параллельно с этим осуществлялись обратные транскрипции. С помощьютеломеразы концы ДНК наращивались теломерными повторами ТТАGGG, роль матрицы при этом выполняла РНК, включающая 450 мононуклеотидов. Поскольку многократно повторенные теломерные последовательности не являются кодирующими, их утрата в процессе деления не приводит к потере информативных участков, тем самым обеспечивается передача генетического материала в поколениях клеток. Однако в родившемся организме активность теломераз регистрируется лишь в половых, раковых, стволовых клетках, в соматических же эти ферменты не работают. Теломеры постепенно укорачиваются, что в конце концов запускает процессы остановки клеточного цикла и провоцирует апоптоз. Интересный факт: ученые, исследующие рак, до сих пор используют культуру клеток Неlа из опухоли женщины по имени Генриетта Лакс. Эта больная умерла в 1951 году, но клетки ее новообразования продолжают делиться и собираются жить вечно. Происходящее в момент деления клетки удлинение синтезирующихся нитей ДНК сопровождается их созреванием, что включает спирализацию, суперспирализацию (закручивание цепей вокруг гистоновых нуклеосом), химическую модификацию (гидроксилирование, метилирование, гидрирование азотистых оснований) с образованием минорных оснований, соединение с помощью ионных связей с белками, катионами металлов (К+, Са++, Мg++, Мn++, Fе++, Сu++и т.д.), которые стабилизируют или, наоборот, дестабилизируют вновь синтезированную молекулу. Синтез и процессинг РНК В отличии от ДНК, синтез которой происходит в момент деления клетки и служит источником генетической информации для последующих поколений, различные РНК необходимы для генерирования молекул белков. Процесс синтеза рибонуклеиновых кислот с использованием в качестве матрицы фрагментов ДНК называют транскрипцией. Все гены в хромосоме (ДНК с белками) в отсутствии факторов транскрипции находится в выключенном (репрессированном) состоянии, потому что нуклеосомы блокируют области инициирования каждого промотора. Ген будет транскрибироваться после вытеснения нуклеосомы и связывания. Та из двух цепей ДНК, на которой пойдет этот процесс, называется кодирующей. Инициация включает раскручивание участка, разрыв водородных связей, что раскрывает азотистые основания транскриптона, в котором выделяют промотор, собственно ген (единицу транскрипции), терминатор. Первая функциональная единица (промотор) служит для с ним РНК-полимеразы, которая объединяет рибонуклеотиды в последовательность, комплементарную кодирующей цепи гена. Момент терминации распознается терминирующим белком – р-фактором. Интересно, что в районе промотора расположены сигнальные последовательности двух типов. Одна из них указывает, где должна начаться транскрипция, а другая определяет, как часто должно происходить это событие. Первый локус называют ТАТА-бокс, второй – СААТ-бокс. Кроме них регуляторная зона включает энхансеры или сайленсеры, изменяющие скорость транскрипции, а также гормончувствительные элементы (ГЧЭ), после взаимодействия с которыми кортикостероиды, андрогены, эстрогены, Т3, сАМФ регулируют экспрессию генов. Практически все первичные транскрипты РНК у эукариот подвергаются сложному процесссингу, т.е. созреванию, начинающемуся в ядре. Будущая иРНК получается после копирования, реакций расщепления, вычленения, легирования, включения дополнительных адениловых нуклеотидов. Ведь свежеполученная РНК содержит неинформативные вставки, считанные с интронов генов, т.е. представляют чередование траслируемых и нетранслируемых участков (гетерогенная ядерная РНК). Процесс удаления интронов и последующего сшивания экзонов получил названиесплайсинг (от англ.tosplice– соединять концы, сшивать). Он требует наличия специальных ферментов, поэтому осуществляется в крупном рибонуклеопротеидном комплексе – тельце, содержащем РНК и белки, необходимые для эффективного сплайсинга. Эта структура называетсясплайсосома. Кроме того к 5'-концу г/яРНК присоединяется 7-метилгуанозинфосфат (кэпирование), который позднее служит стартовой точкой синтеза белка, к 3'-концу надстраивается полиаденилатный хвост (около 200 мононуклеотидов), защищающий иРНК от действия нуклеаз. Параллельно иРНК уже вышедшая из ядра, плотно упаковывается в пространстве. Первичный транскрипт рРНК не содержит интронов, интенсивно метилируется и расщепляется в ядрышке специфическими РНК-азами на 5S, 18S, 28-рРНК, затем они связываются с белками, образуя рибосомы. Подобным образом (путем частичного гидролиза) осуществляется из первичных транскриптов образование тРНК, формирование вторичной, третичной структуры, химическая модификация. Критическое значение имеет этап сплайсинга в области антикодона, т.к. от него зависит точность выполнения адапторной функции при синтезе белка. |
Последнее изменение этой страницы: 2019-04-19; Просмотров: 234; Нарушение авторского права страницы

 ежащего:
ежащего: