|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
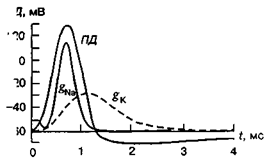
Зміни збудливості клітини під час збудження.
Під час натрієвої інактивації будь-яка деполяризація мембрани абсолютно неефективна, тобто клітина в цей час незбудлива. Це абсолютний рефрактерний період, який триває 1—2 мс. Потім протягом кількох мілісекунд після закінчення абсолютної рефрактерності нове збудження можна викликати лише більшою силою подразнення, ніж перший ПД. Значення ПД в цей час також зменшене, оскільки натрієва система не повністю відновлена після інактивації під час першого ПД. Цей відрізок часу, протягом якого відбувається відновлення збудливості клітини, називають відносним рефрактерним періодом . Абсолютний рефрактерний період обмежує максимальну частоту генерації ПД живою клітиною. Оскільки у більшості нервових клітин П Д триває близько 2 мс, то максимально можлива частота генерації ПД у них буде 500 за 1 с. Проте є клітини з ще коротшим рефрактерним періодом, які мають частоту генерації до 1000 за 1 с. Зазвичай частота генерації ПД в ЦНС не перевищує кількох десятків ПД за 1 с.
Рис. Мембранна провідність під час розвитку ПД у гігантському аксоні кальмара
Під час слідової деполяризації збудливість клітини підвищується (фаза екзальтації), а під час слідової гіперполяризації — знижується (фаза субнормальності), оскільки значення МПС у цей час зростає, при цьому збільшується і критичний (пороговий) рівень деполяризації мембрани.
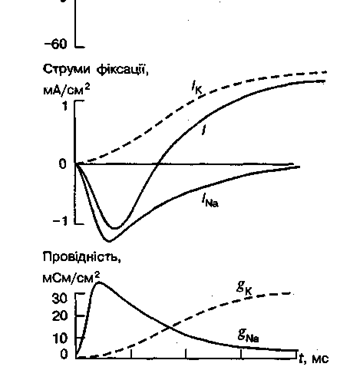
Іонні канали У живій клітині існують дві системи руху іонів крізь мембрану. Одна з них здійснюється за градієнтом концентрації іонів і не потребує витрати енергії — це пасивний іонний транспорт. Він відповідає за підтримання МПС і виникнення ПД, здійснюється через іонні канали. Друга система руху іонів крізь мембрану відбувається проти концентраційного градієнта і полягає у " викачуванні" Na+ з цитоплазми і " закачуванні" К+ всередину клітини за допомогою і онних насосів. Цей механізм можливий тільки з витратами енергії і називається активним іонним транспортом . Функції іонних каналів. У клітинній мембрані існують пори, або канали. Терміном " канал" позначають той шлях, яким іони проходять крізь мембрану за електрохімічним градієнтом. У клітинній мембрані існує кілька різних іонних каналів: селективних та неселективних . Перші пропускають лише якийсь певний іон: Na+, К+ або Са2+, а другі — кілька іонів. Крім того, селективні канали поділяють на потенціалозалежні, потенціалонезалежні (хемочутливі) і канали витоку. Потенціалозалежні канали. Провідність цих каналів контролюється рівнем МПС, тобто ці канали відкриваються (активуються) і закриваються (інактивуються) у відповідь на зміни МПС. Потенціалозалежні канали функціонують в електрозбудливих мембранах і беруть участь у генерації ПД. Електрична збудливість пов’язана з існуванням у таких каналах особливого молекулярного пристрою — воріт, відкривання і закривання яких визначається електричним полем мембрани. Деполяризація мембрани, мВ
Мал. Іонні струми і зміна провідності при зміні МП у гігантському аксоні кальмара: зверху показано деполяризаційне зрушення потенціалу від -60 до 0 мВ. Верхні криві показують підсумковий струм фіксації I та його компоненти Iк і INa; нижні криві показують часовий перебіг змін мембранної провідності gNа і gк, розрахованих за рівнем струмів фіксації
Ворота можуть перебувати тільки у двох положеннях: повністю відчиненими чи повністю зачиненими, тому провідність поодинокого іонного каналу є сталою. Через відкриті канали іони рухаються за концентраційним і електрохімічним градієнтами. Ці потоки іонів зумовлюють зміну МПС, що, в свою чергу, змінює середню кількість відкритих каналів і відповідно рівень іонних струмів. Для дослідження функції іонних каналів застосовують різні методи. Один із них це метод фіксації напруги на мембрані за допомогою якого на мембрані штучно підтримується будь-який потрібний потенціал. Функцію каналів відбивають іонні струми, які в цей час проходять крізь мембрану. Потенціалозалежними є натрієві, калієві і кальцієві канали. Натрієві і кальцієві канали відповідають за ранній (початковий) струм вхідного (в клітину) напрямку, а калієві — за пізній (відставлений) струм вихідного (з клітини) напрямку. Рівень gNа і gк показує кількість одночасно відкритих натрієвих і калієвих каналів. Як можна побачити, gNа за частки мілісекунди досягає максимуму, а потім повільно знижується до початкового рівня. Це пояснюється наявністю у натрієвих каналах двох типів воріт — швидких активаційних і повільних інактиваційних . Отже, початкове збільшення gNа пов'язане з відкриванням перших воріт (процес активації), а наступне повільне зниження gNа, що відбувається під час деполяризації мембрани, зумовлене: закриванням других воріт (процес інактктивації). Натрієвим каналам властива досить висока, але не абсолютна вибірковість Nа+, оскільки більшою або меншою вони є проникними також для іонів літію, талію і навіть деяких органічних катіонів (гідразин, амоній). Отвір натрієвого каналу має прямокутну форму розмірами 0, 3х0, 3 нм. Вузька частина каналу, що є селективним фільтром, дуже коротка і має одиничний негативний заряд. Цей заряд притягує катіони, відштовхує аніони, забезпечуючи вибіркову проникність для Nа+ і подібних до нього катіонів.
а б в Мал. Потенціалозалежний натрієвий канал: Внутрішня частина каналу (1), утвореного макромолекулою білка (2), — це селективний фільтр; т — активаційні. h — інактиваційні: - ворота каналу; а — стан спокою (активаційні ворота зачинені, інактнваційні — відчинені); б — початок деполяризації мембрани (швидке відчинення активаційних воріт); в — інактивація каналу (інактиваційні ворота зачинені); поверхня клітинної мембрани: А — зовнішня; Б — цитоплазматична
Активаційні та інактиваційні ворота розміщені біля внутрішнього кінця натрієвого каналу, причому інактиваційні ворота спрямовані в бік цитоплазми. У стані спокою активаційні ворота закриті, а інактиваційні відкриті. Під час деполяризації мембрани спочатку відкриті обидва типи воріт, тобто канал пропускає Nа+. Потім інактиваційні ворота повільно закриваються — канал інактивується. Лише після закінчення деполяризації поступово відкриваються іиактиваційні ворота, а активаційні закриваються і канал повертається до початкового стану. Деякі речовини специфічно змінюють процеси активації та інактивації натрієвих каналів. Так, батрахотоксин спричинює стійку деполяризацію клітинної мембрани, підвищуючи gNa. Він повністю усуває процес натрієвої інактивації так, що натрієві канали стають постійно відкритими. Тетродотаксин дуже швидко і різко пригнічує ранній іонний струм крізь мембрану, але тільки коли він діє із зовні, тобто на чутливі до тетродотоксину рецепторні структури натрієвих каналів, розміщені на зовнішній поверхні мембрани. Калієві канали за будовою подібні до натрієвих, але відрізняються своєю вибірковістю, спрямованістю потоку іонів, кінетикою процесів інактивації і активації. В усіх збудливих клітинах калієві канали відповідають за пізній іонний струм. Діаметр калієвого каналу у найвужчій його частині становить 0, 3 нм. Отже, калієві канали проникні для катіонів розміром 0, 26-0, З нм (К+, RЬ+). Активація цих каналів порівняно з натрієвими відбувається досить повільно. Крім того, протягом перших 10 мс деполяризації немає ніякої калієвої інактивації, вона виникає тільки під час дуже тривалої (кілька секунд) деполяризації мембрани. Викладене вище про співвідношення між процесами активації та інактивації калієвих каналів властиве лише нервовим волокнам. У мембрані багатьох нервових і м'язових клітин існують калієві канали, які інактивуються порівняно швидко. Виявлено також калієві канали з швидкою активацією. Нарешті, є калієві канали, які активуються не мембранним потенціалом, а внутрішньоклітинним Са2+. Щільність розміщення калієвих каналів на мембрані дещо менша, ніж натрієвих. Специфічним блокатором калієвих каналів є тетраетиламоній (ТЕА), і они гідрогену, амінопіридини. ТЕА діє з будь-якого боку клітинної мембрани. Кальцієві канали. Ретельне дослідження функції іонних каналів різних нервових клітин дало змогу модифікувати іонну модель генерації ПД, запропоновану Ходжкіним і Хакслі, до якої входили лише натрієві й калієві канали. Виявилося, що Са2+ також беруть активну участь у генерації ПД. З'ясувалося також, що в деяких клітинах вхідний струм створюється переважно Са2+ (гладком'язові клітини). Крім порівняно швидкого вхідного кальцієвого струму, що бере участь у генерації ПД, було виявлено ще один дуже повільний кальцієвий струм. Цей струм відповідає за повільну деполяризацію (протягом кількох секунд), яка зумовлює генерацію серій імпульсів деяких пейсмекерних клітин, наприклад серцевих провідних міоцитів (клітин Пуркіньє). Селективність (вибірковість) кальцієвих каналів зумовлена наявністю в ділянці зовнішнього входу хімічних груп, які мають підвищену спорідненість до двовалентних катіонів: іони кальцію зв'язуються з такими групами і тільки після цього проходять до порожнини каналу. До деяких двовалентних катіонів, наприклад Мn2+, спорідненість цих хімічних груп така велика, що, зв'язуючись з ними, ці катіони блокують рух Са2+ через канал. Характерною особливістю кальцієвих каналів є залежність їх від клітинного метаболізму, зокрема від циклічних нуклеотидів (цАМФ, цГМФ), які регулюють процеси фосфоритування і дефосфоритування білків кальцієвих каналів. Іони кальцію ефективно впливають на синтез білків і забезпечують транспорт утворених молекул до аксонів і дендритів. Відкриття цього явища безпосередньо свідчило про те, що процеси на мембрані прямо пов'язані з процесами всередині клітини. Дослідження останніх років показали, що Са2+ є важливим регулятором активності збудливих систем. Переважна кількість Са2+, що потрапляє в клітини під час збудження, зв'язується буферними системами цитозолю, мітохондріями та іншими внутрішньоклітинними депо. Проте та невелика частина Са2*, що залишається вільною, бере активну участь у таких процесах, як регуляція функції К+ і Сl— каналів, Са2+, Nа+ обміну тощо. Виявлено ще одну особливість кальцієвих каналів: вони здатні дуже специфічно реагувати на деякі фізіологічні речовини, що утворюються іншими клітинами і виділяються у внутрішнє середовище організму (катехоламіни, пептиди тощо). Під впливом цих речовин канали активуються чи, навпаки, інактивуються Внаслідок цього змінюється активність нервової клітини. Потенціалонезалежні (хемочутливі) канали. Активність цих каналів контролюється тільки хеморецепторами. Активація хеморецепторів медіаторами супроводжується здебільшого відкриванням каналів. Хемочутливі потенціалонезалежні канали функціонують у субсінаптичних мембранах і беруть участь у: генерації синоптичних потенціалів. Вони реагують не тільки на нейромедіатори, а й на гормони та деякі інші фізіологічно активні речовини. Канали витоку також потенціалонезалежні, але вони не контролюються хеморецепторами. Ці канали відіграють істотну роль у сумарній провідності мембран у стані спокою, саме їхньою активністю пояснюють менші значення МПС порівняно з розрахованим за рівнянням. Функцію цих каналів ще не з’ясовано. Кожний потенціалозалежний іонний канал навіть за сталих значень МПС то відкривається, то закривається. Це відбувається незакономірно, тобто процес має ймовірний (стохастичний) характер. Таке явище викликає переміщення іонів і створює електричний шум. Аналіз цих шумів дав змогу розрахувати щільність розміщення іонних каналів у мембрані і провідність одного каналу. Так, у гігантському аксоні кальмара щільність натрієвих каналів становить 300 на 1 мкм2, середня провідність –4 пСм (пікосименсів); у вузлах нервового волокна(перетяжках Ранв'є) щільність натрієвих каналів значно вища — 2000 на 1 мкм; а провідність удвічі більша — 8 пСм. _
Біоструми Виходячи з усього вищевикладеного, ви можете легко зрозуміти відкіля беруться біоструми в живій тканині і чому вони відсутні в мертвих клітинах. Історія вчення про біоструми нараховує вже декілька сторіч. У 1791 році італійський фізіолог Луіджі Гальвані довів, що в живій тканині є біоструми. Наявність біострумів він довів дослідами, які ввійшли в історію за назвою 1 і 2 дослідів Гальвані. Він установив наявність, так називаних, токів покою або токів ушкодження. Крім того, під час збудження на поверхні мембрани реєструються струми дії. Між ними є розходження. Якщо струми ушкодження однофазні і реєструються довгостроково, то токи дії двофазні і короткочасні. Лабільність У зв'язку зі зміною потенціалу дії, а отже і з тривалістю проходження фаз збуджуваності, знаходиться і функціональна рухливість або лабільність тканини. Вчення про лабільність створив російський учений Н. Е. Введенський. Для визначення частоти виникаючих потенціалів дії, він у 1884 році застосував слухавку, сполучену з нервом нервово - м'язового препарату. При подразненні нерва з різноманітною частотою в слухавці виникали звуки - тони різноманітної висоти. Чим частіше завдавалося подразнення, тим вище був тон. По граничній висоті тону можна було судити про максимальну кількість відтворених у секунду імпульсів збудження. Таке ж положення спостерігалося і при подразненні м'язів. Н. Е. Введенський створив вчення про функціональну рухливість, або лабільність, що розкриває закономірності ритмічного збудження, що поширюється Основним у вченні про лабільність є положення про те, що в кожній тканині одиночний імпульс збудження продовжується визначений час. |
Последнее изменение этой страницы: 2019-06-08; Просмотров: 373; Нарушение авторского права страницы