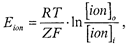|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Роль проницаемости клеточной мембраны и ее поверхностных зарядов
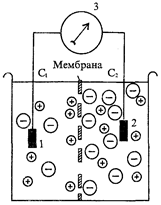
А. Терминология. В настоящее время различные авторы по-разному трактуют термины «проницаемость» и «проводимость». Под проницаемостью клеточной мембраны мы понимаем ее способность пропускать воду и частицы - заряженные (ионы) и незаряженные согласно законам диффузии и фильтрации. Проницаемость клеточной мембраны определяется следующими факторами: 1) наличием в составе мембраны различных ионных каналов -управляемых (с воротным механизмом) и неуправляемых (каналы утечки); 2) размерами каналов и размерами частиц; 3) растворимостью частиц в мембране (клеточная мембрана проницаема для растворимых в ней липидов и непроницаема для пептидов). Термин «проводимость» следует использовать только применительно к заряженным частицам. Следовательно, под проводимостью мы понимаем способность заряженных частиц (ионов) проходить через клеточную мембрану согласно электрохимическому градиенту (совокупность электрического и концентрационного градиентов). Как известно, ионы, подобно незаряженным частицам, переходят через мембрану из области с высокой концентрацией в область с низкой концентрацией. При большом градиенте концентрации и хорошей проницаемости мембраны, разделяющей соответствующие растворы, проводимость ионов может быть высокая, при этом наблюдается односторонний ток ионов. Когда концентрация ионов по обе стороны мембраны уравняется, проводимость ионов уменьшится, односторонний ток ионов прекратится, хотя проницаемость сохранится прежней - высокой. Кроме того, проводимость иона при неизменной проницаемости мембраны зависит и от заряда иона: одноименные заряды отталкиваются, разноименные притягиваются, т.е. важную роль в проводимости иона играет его электрический заряд. Возможна ситуация, когда при хорошей проницаемости мембраны проводимость ионов через мембрану оказывается низкой или нулевой, - в случае отсутствия движущей силы (концентрационного и /или электрического градиентов). Таким образом, проводимость иона зависит от его электрохимического градиента и от проницаемости мембраны: чем они больше, тем лучше проводимость иона через мембрану. Перемещения ионов в клетку и из клетки согласно концентрационному и электрическому градиентам в состоянии покоя клетки осуществляются преимущественно через неуправляемые (без воротного механизма) каналы (каналы утечки). Неуправляемые каналы всегда открыты, они практически не меняют своей пропускной способности при электрическом воздействии на клеточную мембрану и ее возбуждении. Неуправляемые каналы подразделяются на ионоселективные каналы (например, калиевые медленные неуправляемые каналы) и иононеселективные каналы. Последние пропускают различные ионы: К+, Na+, C1-. Б. Роль проницаемости клеточной мембраны и различных ионов в формировании ПП (рис. 3.2.). Сосуд разделен полупроницаемой мембраной. Обе его половины заполнены раствором K2SО4 различной концентрации (C1 и C2), причем C1 < С2. Мембрана проницаема для иона К+ и непроницаема для SО42. Ионы К+ перемещаются согласно концентрационному градиенту из раствора С2 в раствор С1. Поскольку ионы SО42 не могут пройти в раствор С1, где их концентрация тоже ниже, мембрана поляризуется и между двумя ее поверхностями возникает разность электрических потенциалов, соответствующая равновесному калиевому потенциалу (Ек).
Рис. 3.2. Модельный опыт, иллюстрирующий роль избирательной проницаемости мембраны для ионов К+ (+) и S04 (-) в формировании ПП. 1, 2 - электроды; 3 - регистратор; С - концентрация
Ионы Na+ и К+ в живой клетке, находящейся в состоянии покоя, также перемещаются через мембрану согласно законам диффузии, при этом К+ из клетки выходит в значительно большем количестве, чем входит Na+ в клетку, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости для Na+. Органические анионы из-за своих больших размеров не могут выходить из клетки, поэтому внутри клетки в состоянии покоя отрицательных ионов оказывается больше, чем положительных. По этой причине клетка изнутри имеет отрицательный заряд. Интересно, что во всех точках клетки отрицательный заряд практически одинаков. Об этом свидетельствует одинаковая величина ПП при введении микроэлектрода на разную глубину внутрь клетки, как это имело место в опытах Ходжкина, Хаксли и Катца. Заряд внутри клетки является отрицательным как абсолютно (в гиалоплазме клетки содержится больше анионов, чем катионов), так и относительно наружной поверхности клеточной мембраны. Калий является главным ионом, обеспечивающим формирование ПП. Об этом свидетельствуют результаты опыта с перфузией внутреннего содержимого гигантского аксона кальмара солевыми растворами. При уменьшении концентрации ионов К+ в перфузате ПП снижается, при увеличении их концентрации ПП повышается. В состоянии покоя клетки устанавливается динамическое равновесие между числом выходящих из клетки и входящих в клетку ионов К+. Электрический и концентрационный градиенты противодействуют друг другу: согласно концентрационному градиенту К+ стремится выйти из клетки, отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому. Когда концентрационный и электрический градиенты уравновесятся, число выходящих из клетки ионов К+ сравнивается с числом входящих ионов К+ в клетку. В этом случае на клеточной мембране устанавливается так называемый равновесный потенциал. Равновесный потенциал для иона можно рассчитать по формуле Нернста. Концентрация положительно заряженного иона, находящегося снаружи, в формуле Нернста записывается в числителе, а иона, находящегося внутри клетки, - в знаменателе. Для отрицательно заряженных ионов расположение противоположное.
где Eion - потенциал, создаваемый данным ионом; R - газовая постоянная (8, 31 Дм); Т - абсолютная температура (273+37°С); Z -валентность иона; F - постоянная Фарадея (9, 65 х 104); [ion] i - концентрация иона внутри клетки inside; [ion] o - концентрация иона во внешней среде клетки outside. Равновесный потенциал иона Na+ у нервных клеток ENa = +55 мВ, иона калия Е Ek = -70 мВ. Вклад Na+ и Cl- в создание ПП. Проницаемость клеточной мембраны в покое для иона Na+ очень низкая, намного ниже, чем для иона К+, тем не менее она имеется, поэтому ионы Na+ согласно концентрационному и электрическому градиентам стремятся и в небольшом количестве проходят внутрь клетки. Это ведет к уменьшению ПП, так как на внешней поверхности - клеточной мембраны суммарное число положительно заряженных ионов уменьшается, хотя и незначительно, а часть отрицательных ионов внутри клетки нейтрализуется входящими в клетку положительно заряженными ионами Na+. Вход иона Na+ внутрь клетки снижает ПП. Влияние Cl- на величину ПП противоположно и зависит от проницаемости клеточной мембраны для ионов Cl-. Дело в том, что ион Cl-, согласно концентрационному градиенту, стремится и проходит в клетку. Препятствует входу иона Cl- в клетку электрический градиент, поскольку заряд внутри клетки отрицательный, как и заряд Cl-. Наступает равновесие сил концентрационного градиента, способствующего входу иона Cl- в клетку, и электрического градиента, препятствующего входу иона Cl- в клетку. Поэтому внутриклеточная концентрация ионов Cl- значительно меньше внеклеточной. При поступлении иона Cl- внутрь клетки число отрицательных зарядов вне клетки несколько уменьшается, а внутри клетки увеличивается: ион Cl- добавляется к крупным, белковой природы анионам, находящимся внутри клетки. Эти анионы из-за своих больших размеров не могут пройти через каналы клеточной мембраны наружу клетки - в интерстиций. Таким образом, ион Cl-, проникая внутрь клетки, увеличивает ПП. Частично, как и вне клетки, ионы Na+ и Cl- внутри клетки нейтрализуют друг друга. Вследствие этого совместное поступление ионов Na+ и Cl- внутрь клетки не сказывается существенно на величине ПП. В. Определенную роль в формировании ПП играют поверхностные заряды самой клеточной мембраны и ионы Са2+ Наружная и внутренняя поверхности клеточной мембраны несут собственные электрические заряды, преимущественно с отрицательным знаком. Это полярные молекулы клеточной мембраны: гликолипиды, фосфюлипиды, гликопротеиды. Фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, снижают ПП. Фиксированные внутренние отрицательные заряды клеточной мембраны, напротив, суммируясь с анионами внутри клетки, увеличивают ПП. Роль ионов Са2+ в формировании ПП заключается в том, что они взаимодействуют с наружными отрицательными фиксированными зарядами мембраны клетки и нейтрализуют их, что ведет к увеличению и стабилизации ПП. Популярное:
|
Последнее изменение этой страницы: 2016-03-22; Просмотров: 1198; Нарушение авторского права страницы