
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Морфология и анатомия растенийСтр 1 из 48Следующая ⇒
Все ныне живущие растения для удобства изучения подразделяют на две группы — низшие и высшие растения. По современным представлениям к низшим растениям относятся водоросли, а к высшим — все остальные. Тело низших растений, в отличие от высших, не дифференцировано, то есть оно не разделено на органы и ткани. Однородное тело низших растений называют таллом, или слоевище. Дифференциация тела растений произошла в связи с их выходом на сушу. Попав в более контрастные условия окружающей среды, растения были вынуждены вырабатывать специальные приспособления для водоснабжения, защиты от высыхания и т.д. Тело растения разделилось на подземную и надземную части, выполняющие разные функции. Разделение функций привело к возникновению специализированных групп клеток — тканей и органов. Органом называют часть растения, имеющую определенное строение и выполняющую определенные функции. У растений различают вегетативные (обеспечивают процессы питания, дыхания, защиты и вегетативного размножения) и генеративные (выполняют функцию полового размножения) органы. Основными вегетативными органами растений являются корень и побег (лист и стебель рассматриваются как части побега). У низших растений половыми органами (гаметангиями) являются антеридии (мужские) и овогонии (женские), у высших споровых — антеридии и архегонии . У высших семенных антеридии редуцированы, а архегонии имеются только у голосеменных. У цветковых растений цветок, плод и семя называют генеративными органами. Глава 1. Особенности строения Растения, как и все живые организмы, имеют клеточное строение. Они могут быть одноклеточными, колониальными и многоклеточными. Клетка одноклеточного растения представляет собой целый организм и выполняет все функции, необходимые для обеспечения жизнедеятельности. Чаще всего оно имеет форму близкую к шаровидной или яйцевидной. Клетки многоклеточных растений очень разнообразны. Они отличаются друг от друга формой, строением, размерами. Это связано с тем, что в многоклеточном организме клетки выполняют различные функции. Многообразие растительных клеток возникает в результате дифференциации однородных клеток зародыша. Размеры клеток большинства растений колеблются в переделах 10-1000 мкм. Форма клеток многоклеточных организмов может быть округлой, эллипсовидной, кубической, цилиндрической, звездчатой и т.д. Все многообразие форм прастительных клеток можно свести к двум основным типам: © паренхимные клетки — клетки, имеющие форму изодиаметрического многогранника, то есть их размеры во всех трех измерениях приблизительно одинаковы; © прозенхимные клетки — сильно вытянутые клетки, длина которых превышает их ширину и толщину в 5 и более раз (например, волокна льна имеют длину 0, 2-4 см, а толщина не превышает 100мкм. Несмотря на разнообразие, клетки растений имеют общий план строения (рис. 1). Растительная клетка имеет все органоиды, свойственные другим эукариотическим организмам (животные, грибы): ядро, эндоплазматическая сеть, рибосомы, митохондрии, аппарат Гольджи и т.д. Вместе с тем, она отличается от них наличием: © прочной клеточной стенки; © пластид; © развитой системы постоянно существующих вакуолей. Кроме того, в клетках большинства высших растений отсутствует клеточный центр с центриолями. Общий план строения эукариотической клетки рассматривается в разделе " Общая биология" В этой главе мы остановимся только на отличительных особенностях строения растительной клетки.
Растительная клетка, как и животная, окружена цитоплазматической мембраной, поверх которой располагается, как правило, толстая клеточная стенка, отсутствующая у животных клеток. Основным компонентом клеточной стенки является целлюлоза (клетчатка). Молекулы целлюлозы собраны в пучки — фибриллы, образующие каркас клеточной стенки. Промежутки между фибриллами заполнены матриксом, в состав которого входят другие полисахариды — гемицеллюлозы, пектины и гликопротеины. Помимо полисахаридов, в клеточной стенке можно обнаружить и неуглеводные компоненты — лигнин, воска, кутин и суберин. Функции клеточной стенки: © придает клетке определенную форму и прочность; © защищает живое содержимое клетки; © играет определенную роль в поглощении, транспорте и выделении веществ; © служит местом накопления некоторых запасных веществ.
Плазмодесмы — цитоплазматические тяжи, соединяющие содержимое соседних клеток. Они проходят через клеточную стенку. Плазмодесмы представляют собой узкие каналы, выстланные плазматической мембраной. В нем располагается десмотрубочка — цилиндрическая трубочка меньшего диаметра, сообщающаяся с ЭПР обеих соседних клеток. Чаще всего плазмодесмы формируются во время клеточного деления.
Двумембранные органеллы, характерные для растительных клеток. Совокупность всех пластид клетки называется пластидом. Образование пластид происходит из пропластид — мелких телец, находящихся в меристематических клетках корней и побегов. По форме пропластиды напоминают митохондрии, отличаясь лишь большими размерами. Снаружи они покрыты двойной цитоплазматической мембраной. В пластидах различают более или менее развитую мембранную систему (часто это одиночные тилакоиды, расположенные без определенной ориентации; иногда — трубочки или пузырьки) и внутреннее содержимое, представленное гомогенным веществом — строму. Различают три основных типа пластид: © лейкопласты — бесцветные пластиды в клетках неокрашенных частей растений; © хромопласты — окрашенные пластиды обычно желтого, красного и оранжевого цвета; © хлоропласты — зеленые пластиды. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Наиболее часто происходит превращение лейкопластов в хлоропласты (позеленение клубней картофеля на свету) обратный процесс происходит в темноте. При пожелтении листьев и покраснении плодов хлоропласты превращаются в хромопласты. Считают невозможным только превращение хромопластов в лейкопласты или хлоропласты.
Основная функция хлоропластов — фотосинтез, т.е. в хлоропластах на свету осуществляется синтез органических веществ из неорганических за счет преобразования солнечной энергии в энергию молекул АТФ. Хлоропласты высших растений имеют размеры 5-10 мкм и по форме напоминают двояковыпуклую линзу. Хлоропласты — двумембранные органоиды (рис. 2). Наружная мембрана гладкая, а внутренняя имеет складчатую структуру. В результате образования выпячиваний внутренней мембраны, возникает система основных структурных элементов хлоропласта — тилакоидов. Различают: © тилакоиды гран, имеющие вид уплощенных мешочков, уложенных в стопки — граны; ©
Молекулы хлорофилла входят в состав мембран тилакоидов гран, где они собраны в группы — квантосомы. Тилакоиды гран связаны друг с другом таким образом, что их полости оказываются непрерывными. В каждом хлоропласте находится в среднем 40-60 гран, расположенных в шахматном порядке. Этим обеспечивается максимальная освещенность каждой граны. Каждая грана содержит ферменты, участвующие в синтезе АТФ. Внутренняя среда хлоропласта — строма — содержит ДНК и рибосомы, благодаря чему хлоропласт способен к автономному делению, как и митохондрии. На рибосомах происходит синтез белков мембран тилакоидов (в том числе и ферментов, осуществляющих световые реакции фотосинтеза). Белки стромы и липиды мембран имеют внепластидное происхождение. Среди белков стромы особое значение имеют белки-ферменты, синтезирующие органические вещества с использованием энергии АТФ
Бесцветные, обычно мелкие пластиды. Встречаются в клетках органов, скрытых от солнечного света — корнях, корневищах, клубнях, семенах. Форма разнообразна — шаровидная, эллипсовидная, гантелевидная, чашевидная и т.д. Тилакоиды развиты слабо. Имеют ДНК, рибосомы, а также ферменты, осуществляющие синтез и гидролиз запасных веществ. Основная функция — синтез и накопление запасных продуктов (в первую очередь крахмала, реже — белков и липидов).
Встречаются в клетках лепестков многих растений, зрелых плодов, реже — корнеплодов, а также в осенних листьях. Содержат пигменты, относящиеся к группе каротиноидов, придающие им красную, желтую и оранжевую окраску. Внутренняя мембранная система отсутствует или представлена одиночными тилакоидами. Значение в обмене веществ до конца не выяснено. По-видимому, большинство из них представляют собой стареющие пластиды. Косвенное биологическое значение состоит в том, что они обусловливают яркую окраску цветков и плодов, привлекающую насекомых-опылителей и других животных для распространения плодов.
Вакуоли представляют собой полости, заполненные клеточным соком и отграниченные от цитоплазмы мембраной, которую называют тонопластом. На долю вакуолей в растительной клетке приходится до 90% ее объема. Причем, вакуоли являются постоянными компонентами растительных клеток в отличие от животных, в которых могут возникать временные вакуоли. Вакуоли развиваются из цистерн ЭПР. В их образовании принимает участие и аппарат Гольджи, в котором упаковываются продукты обмена веществ и затем в виде пузырьков транспортируются в вакуоль. Молодые клетки, как правило, содержат большое количество мелких вакуолей, которые, постепенно сливаясь, образуют одну большую, занимающую практически всю полость клетки. При этом цитоплазма с органоидами и ядро оказываются оттесненными к цитоплазматической мембране, то есть занимают пристенное положение. Клеточный сок, содержащийся в вакуолях, представляет собой слабоконцентрированный водный раствор органических и неорганических веществ, образующих истинные и коллоидные растворы. В вакуолях происходит накопление как запасных веществ, так и конечных продуктов обмена веществ. Кроме того, в вакуолях часто содержатся особые пигменты из группы антоцианов, придающие растительным клеткам голубую, фиолетовую, пурпурную, темно-красную и пунцовую окраску. Функции вакуолей: © накапливают питательные вещества; © изолируют конечные продукты обмена веществ; © поддерживают тургорное давление; © регулируют водно-солевой обмен; © способствуют растяжению и росту клеток; © окрашивают определенные части растений, привлекая опылителей и распространителей плодов и семян; © могут выполнять функцию лизосом. Глава 2. Растительные ткани Ткань — группа сходных по происхождению и строению клеток и неклеточных структур, образующих структурно-функциональный комплекс и выполняющих одинаковые функции. Обычно при классификации учитывают функции, структуру, происхождение и местоположение тканей. Различают шесть основных групп (систем) тканей: © Система меристематических (образовательных) тканей: ¨ апикальная меристема; ¨ латеральная меристема; ¨ интеркалярная меристема; ¨ раневая меристема. © Система покровных (пограничных) тканей: ¨ эпидерма; ¨ перидерма (пробка); ¨ корка (ритидом); ¨ эпиблема. © Система основных тканей: ¨ ассимиляционная (хлорофиллоносная) паренхима (хлоренхима); ¨ запасающая паренхима. © Система механических (арматурных) тканей: ¨ колленхима; ¨ склеренхима. © Система проводящих тканей (сложные ткани, основу которых составляют проводящие элементы): ¨ ксилема; ¨ Флоэма. © Система выделительных (секреторных) тканей: ¨ наружные секреторные структуры; ¨ внутренние секреторные структуры. Образовательные ткани (меристемы) Растения обладают неограниченным ростом благодаря наличию образовательных тканей. Они образованы недифференцированными (паренхимными) округлыми или многогранными клетками без межклетников. Клеточные стенки тонкие, легко растяжимые, цитоплазма густая, вязкая, ядро крупное, занимает центральное положение. Клетки образовательных тканей способны быстро делиться, поэтому они содержат много рибосом и митохондрий. По происхождению различают: ©
© Вторичные меристемы. Возникают на базе первичных. Обеспечивают рост органов преимущественно в ширину. По местоположению различают: © Верхушечные (апикальные) меристемы. Находятся на концах главных и боковых осей стебля и корня, определяют главным образом рост органа в длину (рис.3). © Боковые (латеральные) меристемы. Возникают за счет деятельности первичных ме- ристем. Как правило, обуславливают утолщение осевых органов. © Вставочные (интеркалярные) меристемы. Участки интенсивно делящихся клеток, расположенные обычно в узлах побегов или в основаниях листовых пластинок. Представляют собой остатки верхушечной меристемы. Когда рост междоузлий или листа прекращается, интеркалярная меристема превращается в постоянные ткани, то есть их деятельность кратковременна. Но иногда эти меристемы могут функционировать достаточно долго (например, у оснований междоузлий хвощей, злаков). © Раневые (травматические) меристемы. Появляются в местах механического разрушения тканей из живых клеток различных паренхимных тканей. Обеспечивают зарастание раны, перекрывают доступ возбудителям болезней. Покровные ткани Покровные ткани являются постоянными образованиями. Возникнув, клетки этих тканей уже не делятся. Как правило, покровными тканями называют ткани, покрывающие тело растения и взаимодействующие с внешней средой. Они защищают внутренние ткани от действия неблагоприятных факторов среды, регулируют газообмен и транспирацию. К собственно покровным тканям относятся:
Покровная ткань зоны всасывания корней называется эпиблемой (ризодермой).
Для газообмена и транспирации в эпидерме имеются специальные образования — устьица (рис. 4). Они представляют собой группу высокоспециализированных клеток. Устьице представляет собой щелевидное отверстие в эпидерме, ограниченное двумя клетками бобовидной формы. Это замыкающие клетки. В отличие от остальных клеток эпидермы они содержат хлоропласты. Стенки замыкающих клеток, обращенные в сторону устьичной щели, утолщены. Клетки эпидермы, окружающие замыкающие, называют побочными или прилегающими. Под устьицем находится газовоздушная камера. Замыкающие и побочные клетки, устьичная щель и га- зовоздушная камера образуют устьичный аппарат. Устьица чаще располагаются на нижней стороне листа.
Защитная функция эпидермы может усиливаться наличием кутикулы и воскового налета. Кутикула представляет собой бесклеточное образование. Оно является продуктом деятельности протопласта и состоит из особого вещества — кутина и воскоподобных веществ. Воскоподобные вещества могут входить в состав кутикулы или располагаться на ее поверхности. Кутикула и восковой налет встречаются на плодах, листьях стеблях, частях цветка. Кутикула и восковой налет непроницаемы для воды и почти непроницаемы для газов.
и феллодермы — пробковой паренхимы. Она сменяет эпидерму, которая постепенно отмирает и слущивается. Закладывается преимущественно в стеблях и корнях. Пробка состоит из правильных радиальных рядов плотно расположенных клеток с опробковшими стенками. Содержимое клетки отмирает. Межклетники отсутствуют. Пробка не проницаема для воды и газов. Для газообмена и транспирации в пробке формируются чечевички.
У большинства деревесных растений пробка заменяется коркой, которую иногда называют третичной покровной тканью. При образовании корки новый слой феллогена и перидермы закладывается в основной ткани, лежащей глубже первой наружной перидермы. Вновь образовавшиеся слои пробки отчленяют к периферии органа не только перидерму, но и часть лежащей под ней паренхимы коры. Так возникает толстое многоклеточное и мертвое образование. Так как корка не может растягиваться, при утолщении ствола она лопается и образуются трещины. Популярное:
|
Последнее изменение этой страницы: 2016-03-16; Просмотров: 1626; Нарушение авторского права страницы
 Рис.1. Строение растительной клетки:
1 — аппарат Гольджи; 2 — свободно расположенные рибосомы; 3 — хлоропласт; 4 — межклеточное пространство; 5 — полирибосомы; 6 — митохондрия: 7 — лизосома; 8 — гранулироанный ЭПР; 9 — гладкий ЭПР; 10 — микротрбочки; 11 — пластиды; 12 — плазмодесмы; 13 — клеточная стенка; 14 — ядрышко; 15 — пора в ядерной оболочке; 16 — наружная цитоплазматическая мембрана; 17 — ядерная оболочка; 18 — гиалоплазма; 19 — тонопласт; 20 — вакуоль; 21 — ядро.
Рис.1. Строение растительной клетки:
1 — аппарат Гольджи; 2 — свободно расположенные рибосомы; 3 — хлоропласт; 4 — межклеточное пространство; 5 — полирибосомы; 6 — митохондрия: 7 — лизосома; 8 — гранулироанный ЭПР; 9 — гладкий ЭПР; 10 — микротрбочки; 11 — пластиды; 12 — плазмодесмы; 13 — клеточная стенка; 14 — ядрышко; 15 — пора в ядерной оболочке; 16 — наружная цитоплазматическая мембрана; 17 — ядерная оболочка; 18 — гиалоплазма; 19 — тонопласт; 20 — вакуоль; 21 — ядро.
 Рис. 2. Схема строения хлоропласта:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — строма; 4 — грана; 5 — тилакоид; 6 — тилакоид стромы; 7 — рибосомы; 8 — ДНК.
Рис. 2. Схема строения хлоропласта:
1 — наружная мембрана; 2 — внутренняя мембрана; 3 — строма; 4 — грана; 5 — тилакоид; 6 — тилакоид стромы; 7 — рибосомы; 8 — ДНК.
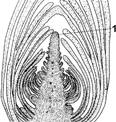 Рис. 3. Конус нарастания:
1 — апикальная меристема конуса нарастания.
Рис. 3. Конус нарастания:
1 — апикальная меристема конуса нарастания.
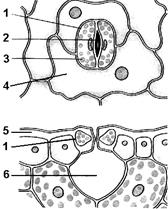 Рис. 4. Строение устьица:
1 — замыкающие клетки; 2 — устьичная щель; 3 — хлоропласты; 4 — прилегающая клетка; 5 — кутикула; 6 — газовоздушная камера.
Рис. 4. Строение устьица:
1 — замыкающие клетки; 2 — устьичная щель; 3 — хлоропласты; 4 — прилегающая клетка; 5 — кутикула; 6 — газовоздушная камера.
 Рис. 5. Перидерма:
1 — чечевичка; 2 — остатки эпидермы; 3 — феллема; 4 — феллоген; 5 — феллодерма.
Рис. 5. Перидерма:
1 — чечевичка; 2 — остатки эпидермы; 3 — феллема; 4 — феллоген; 5 — феллодерма.