|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Картирование генома человека
По данным последнего релиза (2009 г) в аутосомах находится 32593 генов, в половых хромосомах – 2003 гена и в митохондриях – 37 генов. Количество белков в человеческом организме примерно в десять раз больше количества генов. По оценкам большинства авторов, их насчитывается более 300 000, а число белок-белковых взаимодействий и вовсе не поддается подсчету. Многие болезни могут быть прослежены до изменений, происходящих на уровне белков. К примеру, известно, что при серповидно-клеточной анемии аномальный белок гемоглобин вызывает изменение формы красных кровяных телец. Аномальные эритроциты имеют серповидную форму. Серповидно-клеточная анемия обусловлена мутацией, приводящей к замене глутаминовой кислоты на валин в бета-цепи гемоглобина (гемоглобин S). Это заболевание является классическим примером приспособительного значения мутаций – гетерозиготы по замене глутаминовой кислоты на валин в бета-цепи гемоглобина более устойчивы к малярии, поэтому частота ее встречаемости выше в географических районах распространения малярийного плазмодия. Часто после синтеза (трансляции), для того чтобы выполнять определенные функции в организме, белки требуют модификации. Причина многих заболеваний человека – модификации белков и изменения характера их взаимодействий. Результаты исследований в области протеомики в последние годы все чаще применяются в биофармакологии, которая получила благодаря этому надежный теоретический фундамент. Сегодня более 95 % всех имеющихся на рынке лекарственных средств оказывают свое действие именно на белки. Системные подходы протеомики помогают гораздо эффективнее идентифицировать и оценивать новые целевые белки, а следовательно, ускорить разработку новых диагностических систем и терапевтических средств и сделать их более эффективными.
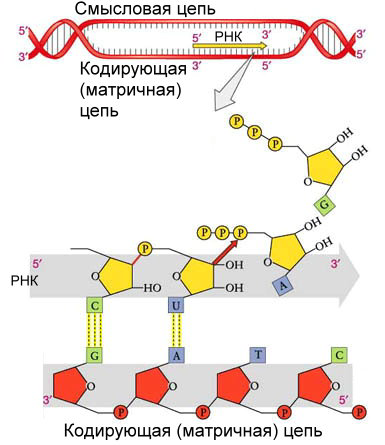
ТЕМА 7. ФУНКЦИОНИРОВАНИЕ КЛЕТКИ Биосинтез белка Одно из наиболее важных и характерных свойств живой клетки. Первичная структура белка, как уже отмечалось, предопределяется генетическим кодом, заложенным в молекуле ДНК, причем различные ее участки кодируют синтез разных белков. Следовательно, одна молекула ДНК хранит информацию о структуре многих белков. Свойства белка зависят от последовательности расположения аминокислот в полипептидной цепи. В свою очередь чередование аминокислот определяется последовательностью нуклеотидов в ДНК. В иРНК каждой аминокислоте соответствует определенный триплет — группа, состоящая из трех нуклеотидов, называемая кодоном. Биосинтез белка начинается в ядре со списывания информации о структуре белковой молекулы с ДНК на иРНК по принципу комплементарности. Данный процесс протекает как реакция матричного синтеза и называется транскрипцией (рис. 7.1).
Рис. 7.1. Процесс транскрипции
В результате транскрипции образуется «незрелая» иРНК (пре-иРНК), которая проходит стадию созревания или процессинга.
Процессинг включает в себя: 1) КЭПирование 5'-конца; 2) полиаденилирование 3'-конца (присоединение нескольких десятков адениловых нуклеотидов); 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиА «хвост». Возможен альтернативный сплайсинг, при котором вместе с интронами вырезаются и экзоны. При этом с одного гена могут образовываться разные белки. Таким образом, утверждение – «Один ген – один полипептид» – неверно (рис. 7.2, 7.3, 7.4)
Рис. 7.2. Сплайсинг
Рис. 7.3. Альтернативный сплайсинг (варианты)
Рис. 7.4. Образование разных молекул белка при вариантах альтернативного сплайсинга
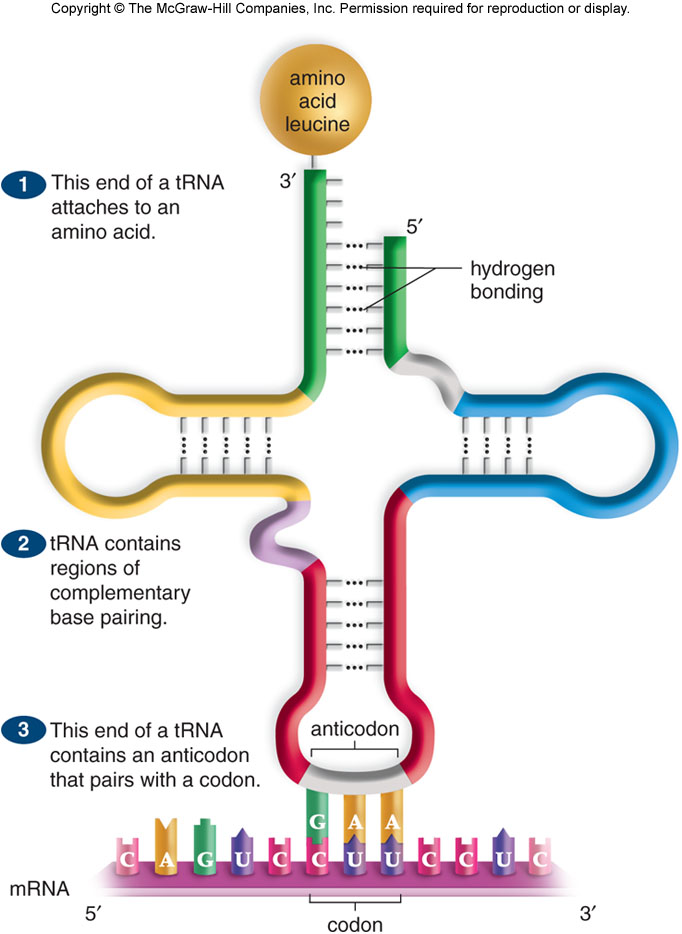
Образующаяся при этом иРНК поступает в цитоплазму, где на нее нанизываются рибосомы. Одновременно в цитоплазме с помощью ферментов активизируется транспортная РНК тРНК. Молекула тРНК напоминает по структуре лист клевера, на вершине которого находится триплет нуклеотидов, соответствующий по коду определенной аминокислоте (антикодон), а основание («черешок») служит местом присоединения этой аминокислоты. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3'-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ) к участку ССА (рис. 5) Транспортная РНК доставляет аминокислоты к рибосомам. По принципу комплементарности антикодон связывается со своим кодоном, причем аминокислота располагается у активного центра рибосомы и с помощью ферментов соединяется с ранее поступившими аминокислотами. Затем тРНК освобождается от аминокислоты, а молекула иРНК продвигается вперед на один триплет, и процесс повторяется.
Рис. 7.5. Строение молекулы Т-РНК
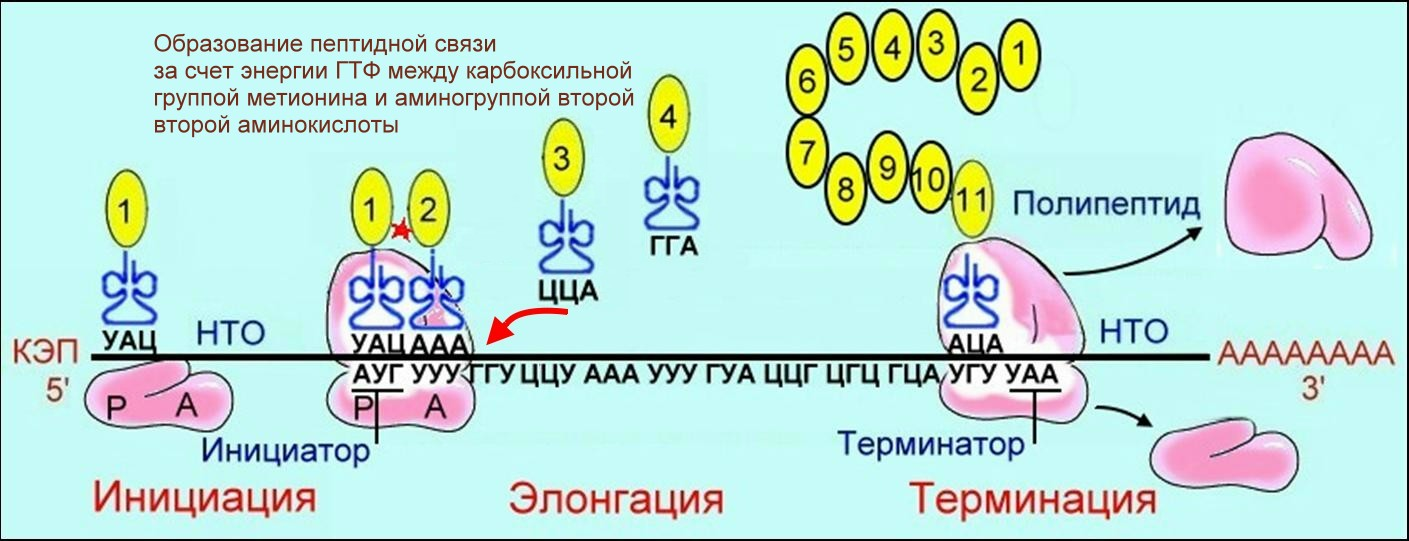
Различают три этапа в биосинтезе белка: инициацию, элонгацию и терминацию. В малой субъединице рибосомы расположен функциональный центр рибосомы (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три - в пептидильном и три - в аминоацильном участках. Инициация. Синтез белка начинается с того момента, когда к 5'-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК. За счет АТФ происходит передвижение инициаторного комплекса (малая субъединица рибосомы, тРНК с метионином) по НТО до метионинового кодона АУГ. Этот процесс называется сканированием. Элонгация. Как только в Р-участок сканирующего комплекса попадает кодон АУГ, происходит присоединение большой субъединицы рибосомы. В А-участок ФЦР поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке. Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислоты. Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Терминация. Когда в А-участок попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения, полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Рис. 7.6. Процесс трансляции (шаг 1)
Рис. 7.7. Процесс трансляции (шаг 2)
Рис. 7.8. Процесс трансляции (шаг 3)
Рис. 7.9. Процесс трансляции (шаг 4)
Рис. 7.10. Биосинтез белка (общая схема) Так постепенно наращивается белковая цепочка, в которой аминокислоты располагаются в строгом соответствии с локализацией кодирующих их триплетов в молекуле иРНК. Синтез полипептидных цепей белков по матрице иРНК называется трансляцией (рис. 10). В клетках растительных и животных организмов белки непрерывно обновляются. Интенсивность синтеза тех или иных специфических белков определяется активностью соответствующих генов, с которых «считывается» иРНК. Следует отметить, что не все гены функционируют одновременно: активность проявляют лишь те, которые кодируют информацию о структуре белков, необходимых для жизнедеятельности организма в данный момент.
Генетический код
Кодирование генетической информации о структуре белка осуществляется посредствам генетического кода. Зависимость между последовательностью оснований ДНК и последовательностью аминокислотных остатков в белке получила название генетического кода. Первым кто предложил абстрактную гипотезу кодирования, а также способ её проверки, был советский и американский физик-теоретик Георгий Гамов (рис. 7.11). В 1954 году Гамов опубликовал свою работу, в которой предложил в качестве механизма кодирования установление соответствия между боковыми цепями аминокислот и ромбовидными «дырами», образованными четырьмя нуклеотидами ДНК. Позднее этот код был назван ромбическим или бубновым. Исходя из своей модели Гамов предположил, что код может быть триплетным. Несмотря на все очевидные недочёты этой гипотезы (например, идея о том, что структура белка напрямую кодируется ДНК) она стала первой среди многих гипотез о природе кода. Гамов был первым, кто представил проблему кодирования.
Рис. 7.11. Георгий Гамов (1904-1968)
За несколько последующих лет было предложено большое количество разных моделей. Все предложенные коды можно разделить на две категории: перекрывающиеся (один нуклеотид входит в состав более чем одного кодона) и неперекрывающиеся. В конце пятидесятых годов два ученых, М. Ниренберг (рис. 7.12) и Г. Маттеи открыли основы генетического кода. К 1965 г. генетический код был полностью расшифрован. Если бы каждый отдельный из нуклеотидов кодировал одну аминокислоту, то белок мог включать только четыре аминокислоты. Если каждая аминокислота кодировалась бы только двумя основаниями, то это позволило бы закодировать 16 аминокислот. Однако лишь код, состоящий из трех оснований, может обеспечить включение в белок всех известных аминокислот. Подобный код содержит 64 различных сочетания из трех нуклеотидов. Доказательства триплетности генетического кода были впервые получены Ф. Криком в экспериментах с фагом Т4. Из 64 кодонов три кодона УАГ, УАА, УГА не кодируют аминокислот и поэтому были названы бессмысленными или нонсенс-кодонами. Позднее оказалось, что они являются терминирующими кодонами (рис. 7.13).
Рис. 7.12. М. Ниренберг (1927-2010)
Свойства генетического кода 1. Генетический код триплетен. Триплет (кодон) — это последовательность трех нуклеотидов, кодирующая одну аминокислоту. 2. Генетический код является вырожденным: данная аминокислота может кодироваться более чем одним кодоном. Вырожденность кода обусловлена тем, что у кодонов, определяющих одну и ту же аминокислоту, первые два основания фиксированы, а третье положение может занимать другое основание. Исключение составляют метионин и триптофан, которые кодируются только одним триплетом. Триплет, соответствующий метионину (АУГ), инициирует считывание и не кодирует аминокислоту, если стоит в начале цепи ДНК. 3. Генетический код однозначен. Каждому кодону соответствует только одна аминокислота, т.е. триплет шифрует только одну аминокислоту. 4. Код не перекрывается, т.е. в последовательности оснований АБВГДЕЖЗИК первые три основания, АБВ, кодируют аминокислоту 1, ГДЕ – аминокислоту 2 и так далее. Если бы код был перекрывающимся, то кислоту 2 могла бы кодировать последовательность ВГД. В коде отсутствуют запятые, т.е. нет знаков, отделяющих один кодон от другого. 5. Генетический код универсален: одни и те же триплеты кодируют одни и те же аминокислоты у всех живых существ на Земле независимо от уровня их организации (вирусы, прокариоты, эукариоты). Существует 3 генетических кода: в ядре действуеьт универсальный генетический код, в пластидах – пластидный, в митохондриях – митохондриальный генетический код. 6. Наличие кодона-инициатора и кодонов-терминаторов. В генетическом коде имеется кодон-инициатор (метионин), с которого начинается синтез любого полипептида.
Из 64 кодовых триплетов 61 кодон — кодирующие, т.е кодируют аминокислоты. Три кодона являются бессмысленными, некодирующими. Они заканчивают синтез полипептида (УАА, УГА, УАГ).
Рис. 7.13. Таблица генетического кода Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 1544; Нарушение авторского права страницы