|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Как происходит Управление движениями?
Сокращая мышцы, мы изменяем, положение частей тела, т.е. производим движение. В двигательной деятельности человека различают непроизвольные движения, происходящие без участия сознания и представляющие собой безусловные рефлексы, либо автоматизированные двигательные навыки, а также произвольные движения – сознательно управляемые целенаправленные действия. Произвольные движение человека сознательно управляются (контролируются) нервной системой посредством механизма сенсорно-двигательной интеграции. Любое движение можно рассматривать как двигательную реакцию на определенный внешний или внутренний стимул.
Что понимается под термином «сенсорно-двигательная интеграция»? Сенсорно-двигательная интеграция – механизм, посредством которого, нервная система управляет (контролируют) произвольные движения человека взаимодействие сенсорного и двигательного отделов нервной системы, в результате чего осуществляется выполнение двигательных реакций процесс последовательного функционирования сенсорного и двигательного отделов нервной системы, чтобы тело прореагировало на сенсорный стимул процесс последовательного взаимодействия сенсорного и двигательного отделов нервной системы, в результате чего осуществляется управление (контроль) движениями человека Сенсорно-двигательная интеграция представляет собой процесс, посредством которого ПНС передает сенсорный импульс ЦНС, где он интерпретируется, после чего посылается в соответствующий двигательный сигнал, направленный на то, чтобы вызвать необходимую двигательную реакцию. Процесс, который называется сенсорно-двигательной интеграцией, показан на рис. 30. Чтобы тело прореагировало на сенсорный стимул, сенсорный и двигательный отделы нервной системы должны функционировать вместе в такой последовательности событий: 1) сенсорные рецепторы принимают сенсорный стимул; 2). сенсорный импульс передается по сенсорным нейронам в ЦНС; 3) ЦНС обрабатывает поступившую сенсорную информацию и определяет наиболее подходящую реакцию на нее; 4) сигналы реакции передаются из ЦНС по двигательным нейронам; 5) двигательный импульс передается мышце и реакция осуществляется. Сенсорный импульс Ощущения и физиологический статус организма определяют сенсорные рецепторы. Импульсы вследствие сенсорного стимулирования передаются через сенсорные нервы в спинной мозг. Достигнув его, они " включают" локальный рефлекс на данном уровне или идут в более высокие участки спинного мозга или в головной мозг. Сенсорные пути к головному мозгу могут прерываться в сенсорных участках ствола мозга, в мозжечке, таламусе или коре головного мозга. На рис. 31 показаны сенсорные рецепторы, а также их нервные пути к спинному мозгу и различным участкам головного мозга.
Рис. 31. Чувствительные рецепторы и пути к спинному и головному мозгу Функции интеграционных центров различны: · сенсорные импульсы, которые прерываются (заканчиваются) в спинном мозгу, здесь и интегрируются. Реакцией обычно является простой двигательный рефлекс, представляющий собой наипростейший тип интеграции; · сенсорные импульсы, заканчивающиеся в нижней части ствола мозга, вызывают подсознательные двигательные реакции более высокого уровня и более сложные, чем рефлексы спинного мозга, контроль при передвижении, пребывании в положении сидя или стоя; · сенсорные импульсы, которые заканчиваются в мозжечке, также участвуют в реализации подсознательного контроля движения. Это центр координации, делающий наши движения более плавными, координируя действия различных сокращающихся мышечных групп. Мозжечок вместе с базальными ядрами головного мозга координирует все тонкие и грубые движения тела. Без контроля со стороны мозжечка все выполняемые движения были бы нескоординированными и неконтролируемыми; · сенсорные сигналы, заканчивающиеся в таламусе, достигают уровня сознания, и человек начинает различать все возможные ощущения; · когда сенсорные сигналы попадают в кору головного мозга, человек может дискретно локализовать сигнал. Основная чувствительная зона коры головного мозга, расположенная в постцентральной извилине (в теменной доле), принимает общие сенсорные импульсы от рецепторов кожи, а также проприорецепторов мышц, сухожилий и суставов. В этом участке имеется “карта” всего тела. Стимулирование в определенном участке распознается, и его точное нахождение сразу же становится известным. Таким образом, эта часть головного мозга постоянно обеспечивает нас информацией обо всем, что нас окружает, и о нашей взаимосвязи с окружающей средой.
Рис. 32. Двигательные пути нервной системы
ДВИГАТЕЛЬНЫЙ КОНТРОЛЬ После поступления в ЦНС сенсорного импульса сразу же возникает реакция двигательного нейрона, независимо от уровня, на котором " остановился" импульс. Контроль скелетных мышц осуществляют импульсы, проводимые двигательными (эфферентными) нейронами, которые берут начало в одном из трех уровней: 1) спинном мозгу; 2) нижних участках головного мозга; 3) двигательном участке коры головного мозга. По мере перемещения уровня осуществления контроля от спинного мозга к двигательной области коры головного мозга увеличивается сложность движений от простых рефлексов к усложненным движениям, выполнение которых требует участия мыслительных процессов. Двигательные реакции более сложных движений, как правило, берут свое начало в двигательной зоне коры головного мозга. Некоторые двигательные пути показаны на рис. 32. Теперь мы можем связать воедино две системы на основании сенсомоторной интеграции. Простейшей ее формой является рефлекс, с него мы и начнем. Рефлекторная деятельность. Рефлексы представляют собой простейшую форму нервной интеграции. Рефлекс представляет собой заранее запрограммированную реакцию. В любой момент при передаче сенсорными нервами особых импульсов ваше тело реагирует мгновенно и одинаково. К тому времени, когда вы осознанно ощутите особый стимул, после того как сенсорные импульсы будут переданы в чувствительную зону коры головного мозга, рефлекторная деятельность, скорее всего, завершится. Вся нервная деятельность протекает очень быстро, однако рефлекс — наиболее быстрый режим реакции, поскольку не требует принятия решения. Возможна лишь одна реакция — нет необходимости рассматривать варианты. Рассмотрим два рефлекса, которые помогают контролировать мышечную функцию: Рассмотрим два рефлекса, которые помогают контролировать мышечную функцию. Первый из них включает особую структуру — нервно-мышечное веретено.
Рис. 33. Схема: Нервно-мышечное веретено Нервно-сухожильное веретено. 1 - сухожильный орган Гольджи; 2 - капсула; 3 - соединительная капсула; 4 - мышечное веретено; 5 - первичное чувствительное окончание (волокна Iа типа); 6 - вторичное чувствительное окончание (волокна II типа); 7 - гамма-эфферентное двигательное волокно к интрафузальным мышечным волокнам; 8 - альфа - эфферентное двигательное волокно к экстрафузальным мышечным волокнам; 9 - экстрафузальные мышечные волокна; 10 - интрафузальные мышечные волокна; 11 - чувствительное волокно; 12 – сухожилие Нервно-мышечные веретена Нервно-мышечные веретена ( рис. 33 ) находятся между обычными волокнами скелетной мышцы, так называемыми экстрафузальными (снаружи веретена) волокнами. Нервно-мышечное веретено состоит из 20 — 24 маленьких особых мышечных волокон, которые называются интрафузальными (внутри веретена), и нервных окончаний (сенсорных и двигательных, связанных с ними). Оболочка соединительной ткани окружает нервно-мышечные веретена и прикрепляется к внутреннему слою экстрафузальных волокон. Интрафузальные волокна контролируются специальными двигательными нейронами — гамма-двигательными нейронами. Альфа-двигательные нейроны контролируют экстрафузальные (обычные) волокна. Центральный участок интрафузального волокна не может сокращаться, так как не содержит или содержит очень мало актиновых и миозиновых филаментов. Следовательно, этот участок может только растягиваться. Поскольку нервно-мышечное веретено прикреплено к зкстрафузальным волокнам, в любой момент, когда растягиваются эти волокна, растягивается и центральный участок веретена. Чувствительные нервные окончания, окутывающие этот центральный участок нервно-мышечного веретена, сообщают в спинной мозг информацию о его растяжении, информируя ЦНС о длине мышцы. В спинном мозгу сенсорный нейрон взаимодействует с альфа-двигательным нейроном, который вызывает рефлекторное мышечное сокращение экстрафузальных волокон, противодействующее дальнейшему растяжению. Гамма-двигательные нейроны возбуждают интрафузальные волокна, предварительно их слегка растягивая. Если центральный участок этих волокон не может сокращаться, то их концы могут. Гамма-двигательные нейроны вызывают незначительное сокращение окончаний интрафузальных волокон, что приводит к незначительному растяжению центрального участка. Это предварительное растяжение обеспечивает высокую чувствительность нервно-мышечного веретена даже к очень незначительному растяжению. Нервно-мышечное веретено способствует нормальному мышечному сокращению. При стимулировании альфа-двигательных нейронов на сокращение экстрафузальных мышечных волокон гамма-двигательные нейроны также оказываются активизированными и возбуждают окончания интрафузальных волокон. Это приводит к растяжению центрального участка нервно-мышечного веретена и направлению сенсорных импульсов в спинной мозг и затем к двигательным нейронам. В ответ мышца сокращается. Таким образом, нервно-мышечные веретена способствуют мышечному сокращению. Информация, поступившая в спинной мозг от сенсорных нейронов, связанных с нервно-мышечными веретенами, не обязательно остается на этом уровне. Импульсы также направляются в более высокие отделы ЦНС. обеспечивая головной мозг информацией о точной длине и степени сокращения мышцы, а также степени их изменений. Эта информация необходима для поддержания мышечного тонуса, позы и выполнения движений. Прежде чем головной мозг " скажет" мышце, что делать, он должен знать, что она делает в настоящий момент. Нервно-сухожильные веретена Нервно-сухожильные веретена представляют собой сенсорные рецепторы, через которые проходит небольшой пучок мышечно-сухожильных волокон. Они расположены проксимально ее места прикрепления сухожильных волокон к мышечным ( рис. 33 ). Около 5 — 25 мышечных волокон, как правило, соединены с каждым нервно-сухожильным веретеном. Если нервно-мышечные веретена следят за длиной мышцы, то эти структуры чувствительны к напряжению в мышечно-сухожильном комплексе и действуют подобно тензиометру. Их чувствительность настолько высока, что они могут реагировать на сокращение отдельного мышечного волокна. По сути, они являются тормозными и выполняют защитную функцию, снижая вероятность травмы. При стимулировании они тормозят сократительные (агонисты) мышцы и возбуждают антагонистические. По мнению некоторых ученых, уменьшение влияния нервно-сухожильных веретен приводит к растормаживанию активных мышц, обеспечивая более мощное их сокращение. Этот механизм объясняет, по крайней мере частично, прирост мышечной силы вследствие тренировок силовой направленности.
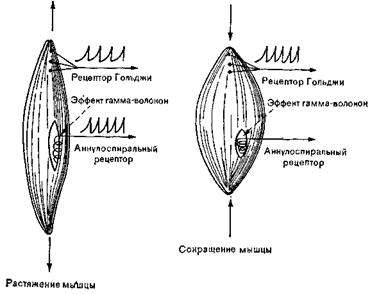
Рис. 34. Схема активации проприоцепторов. Нервно-сухожильные и нервно-мышечные веретена представляют собой сенсорные рецепторы – классические представители проприоцепторов ( рис. 34 ). Аннулоспиральный рецептор - нервно-мышечное веретено - разряжается только при растяжении мышцы, а рецептор Гольджи - нервно-сухожильное веретено - с более высоким порогом, отвечает и на растяжение, и на сокращение Механизм гамма-эфферентов контролирует только аннулоспиральные рецепторы.
ВЫСШИЕ ЦЕНТРЫ ГОЛОВНОГО МОЗГА
Высший уровень сенсорно-двигательной (сенсомоторной) интеграции: • Двигательная область коры головного мозга • Базальные ядра • Мозжечок Выясним некоторые функции каждого из них. Двигательная область коры головного мозга осуществляет контроль тонких и разрозненных мышечных движений. Она находится в лобной доле в передней центральной извилине. Находящиеся здесь нейроны, так называемые пирамидальные, позволяют нам сознательно контролировать движения скелетных мышц. Двигательная область коры головного мозга представляет собой часть головного мозга, которая решает, какое движение вы хотите выполнить. Участки, требующие тончайшего двигательного контроля, в большей степени представлены на ней, тем самым обеспечивается больший нервный контроль. Тела пирамидных нейронов находятся в двигательной области коры головного мозга, и их аксоны образуют экстрапирамидные пути. Это так называемые корково-спинномозговые пути, поскольку нервные процессы проходят от коры головного мозга до спинного мозга без переключения. Эти пути обеспечивают главный произвольный контроль скелетных мышц. Базальные ядра. Базальные ядра не являются частью коры головного мозга. Они находятся в белом веществе, глубоко под корой головного мозга. Базальные ядра представляют собой скопления нейронов. Сложные функции их изучены недостаточно; известна их важная роль в инициации движений продолжительного или повторяющегося характера (таких, как движения рук при ходьбе), следовательно, они контролируют сложные полупроизвольные движения, например, ходьбу и бег. Кроме того, они участвуют в поддержании мышечного тонуса и позы. Мозжечок. Мозжечок играет важнейшую роль в контроле всех быстрых и сложных видов мышечной деятельности. Он помогает синхронизировать двигательную деятельность и быстрый переход от одного движения к другому, следя и внося необходимые изменения в двигательную деятельность, вызванную другими участками головного мозга. Мозжечок содействует функционированию как двигательной области коры головного мозга, таки базальных ядер. Он “смягчает” структуру движений, в противном случае они бы были резкими и не координируемыми. Мозжечок выполняет роль интеграционной системы, сравнивая запрограммированную деятельность с изменениями, которые происходят в организме, и производя затем соответствующие корригирующие действия с помощью двигательного отдела. Он получает информацию из головного мозга, а также от проприорецепторов, находящихся в мышцах и суставах, которые сообщают о положении, занимаемом в данный момент телом. Кроме того, мозжечок получает зрительные импульсы, а также импульсы о равновесии. Следовательно, он обрабатывает всю поступающую информацию о напряжении и положении всех мышц, суставов и сухожилий, а также положении тела относительно окружающих условий и затем определяет наилучший план действий, направленный на выполнение необходимого движения. Таким образом, в мозжечке формируется и при необходимости корректируется программа выполнения произвольного движения. Двигательная программа (энграмма). При освоении любого движения - новой двигательной программы - в первое время необходима высокая концентрация внимания. По мере изучения движения необходимость значительной концентрации внимания снижается. Наконец, когда вы овладеваете этим действием, вы можете произвести его практически без сознательного усилия. Как это достигается? Выработанные структуры движений “хранятся” в мозгу и при необходимости могут быть воспроизведены. Эти структуры, называющиеся двигательными программами, или энграммами, хранятся в сенсорном и двигательном отделах мозга. В сенсорном отделе хранятся структуры более медленных движений, двигательном — быстрых. В настоящее время сведений об энграммах и о механизмах их действия недостаточно. Энграмма — (греч. έ ν — пребывание в каком-либо состоянии или действии + греч. γ ρ ά μ μ α - все написанное, запись) - «внутренняя запись». - англ. engram) - совокупность изменений в нервной ткани, обеспечивающих сохранение результатов воздействия действительности на человека. Э. - физиологическая основа памяти. Два вида энграмм: образы (отображение статической структуры объекта) и модели действий (программы). Двигательная программа (энграмма): запечатленные двигательные программы, которые хранятся в сенсорном и двигательном отделах головного мозга и могут быть воспроизведены при необходимости. В результате активации генома и синтеза специфических белков в первую очередь в мембранах нервных клеток в процессе обучения формируется структурно-функциональное объединение нейронов различных структур мозга, представляющее собой энграмму памяти. С позиций теории функциональных систем такая энграмма в первую очередь строится на структурной основе аппарата акцептора результата действия. Энграмма представляет ансамбль нейрональных и глиальных элементов, объединенных синаптическими механизмами. Такое динамическое объединение строится за счет экспрессии геномом отдельных клеток определенных белковых молекул — адгезинов, или коннектинов, которые встраиваются в специальные области мембран нейронов. Именно эти идентичные по молекулярным свойствам белковые молекулы увеличивают чувствительность нейронов к приему той или иной информации, которая первично вызывала экспрессию этих белков. Вопрос о структурной организации энграммы до сих пор остается открытым. Нейропсихологи проводят исследования в поисках локализации энграммы памяти. Это позволило ряду авторов сформулировать голографическую гипотезу памяти. Много гипотез и мало реальных открытий. Упорядоченное рекруитирование мышечных волокон. Большинство ученых считают, что двигательная (нервно-мышечная) активность дифференцируется на основании фиксированного упорядоченного рекруитирования двигательных единиц. Чем больше сила, необходимая для выполнения определенного движения, тем больше вовлекается двигательных единиц; при выполнении определенной (однотипной) деятельности всегда рекруитируются одни и те же двигательные единицы. Упорядоченное рекруитирование мышечных волокон: принцип размера; кодирование нервной стимуляции движений Что такое принцип размера? Механизм, который хотя бы частично позволяет объяснить это упорядоченное рекруитирование, основан на принципе размера, согласно которому вовлечение двигательной единицы непосредственно связано с размером двигательного нейрона. Первыми рекруитируются двигательные единицы, имеющие небольшие двигательные нейроны. Поскольку медленносокращающиеся двигательные единицы имеют небольшие двигательные нейроны, они рекруитируются первыми для выполнения дифференцированного движения (от низкой до очень высокой интенсивности производства усилия). Затем, с увеличением силы, необходимой для выполнения движения, рекруитируются быстросокращающиеся двигательные единицы. Это связано с тем, что подключение мышечных волокон к работе зависит от силы стимуляции мотонейроном. Минимальная частота стимуляции, при которой волокно сокращается максимально, называется порогом возбуждения (раздражения). Минимальный порог возбуждения имеют медленно сокращающиеся волокна (10-15 Гц); у быстро сокращающихся волокон порог возбуждения в 2 раза выше, чем у медленно сокращающихся волокон. Все типы мышц вовлекаются в работу при высокой частоте раздражения – около 45-55 Гц. Что такое кодирование нервной стимуляции движений? Кодирование нервной стимуляции обеспечивает различные варианты сокращения скелетной мышцы, и, следовательно, различные типы движений. Центральная нервная система кодирует движение посредством выбора наиболее подходящей двигательной единицы, подключения дополнительных двигательных единиц для увеличения силы сокращения (пространственная суммация), а также дополнительной стимуляции одних и тех же двигательных единиц (временная суммация). Кодирование дает возможность одной и той же мышце выполнять как тонкие и точные, так и мощные и грубые движения. Многим исследователям не совсем понятно, как можно применить данные принципы к большинству спортивных движений, поскольку он рассматривается только при выполнении дифференцированных движений, представляющих относительную интенсивность мышечного сокращения менее 25 %. При выполнении многих движений, например, спринт или рывок в тяжелой атлетике, необходимо почти мгновенное включение преимущественно или только быстрых мышечных волокон. Исследователи активно занимаются построением теоретических моделей развития основных пространственно-временных форм траектории движения на основе сложных математических методологий. Это необходимо не только в физиологии спорта высших достижений, но и в современной неврологии для разработки методов восстановления мышечной активности после перенесенных нарушений мозгового кровообращения… процесс продолжается и ожидается много новых интересных находок и достижений.
Заключение Подводя предварительные итоги, нельзя не заметить значительные достижения в изучении физиологии физической активности. Прежде всего, на микро – уровне: хорошо изучены сложные механизмы сокращения мышечного волокна и особенности энергообеспечения этого процесса. Но на макро – уровне (на уровне целого организма) – пока преобладают интересные теории над реальными достижениями. Но растущие потребности медицины (особенно в кардиологии и неврологии) и спорта высших достижений активно продвигают научные исследования в этой сфере. Вопросы для самостоятельного контроля 1. Физическая нагрузка. Физическая активность. Цели и задачи физиологии физической нагрузки 2. Скелетная мышца. Характеристика структурной единицы мышечной ткани скелетной мышцы. Саркомер. Взаимодействие актина и миозина. Функция кальция в скелетных мышцах. Теория скользящих нитей 3. Сопряжение возбуждения и сокращения скелетной мышцы (электромеханическое сопряжение).Функции АТФ в процессе сокращения скелетной мышцы. Роль АТФ в цикле поперечных мостиков. 4. Основные пути ресинтеза АТФ в мышечных волокнах. Дефицит кислорода. Кислородный долг. 5. Характеристика (скорость укорочения и тип метаболизма) волокон скелетных мышц разного типа (1 а, 11а, 11б). 6. Характеристика энергетического пути, используемого во время кратковременной интенсивной физической нагрузки. 7. Характеристика энергетического пути, используемого во время продолжительной физической нагрузки. 8. Двигательная единица. Понятия: " коэффициент иннервации", " низкий коэффициент иннервации", " высокий коэффициент иннервации". Понятие термина: «сенсорно-двигательная интеграция». 9. Роль проприоцепторов нервно-мышечного веретена и нервно-сухожильного веретена в регуляции движения, механизмы их активация. 10. Центры головного мозга, которые обеспечивают высший уровень сенсорно-двигательной (сенсомоторной) интеграции.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА 1. Физиология человека: В 3 томах. Пер с англ./ Под ред. Р.Шмидта и Г.Тевса. – М.: Мир. 1996. Т1 Глава 4. Мышцы – С.69 – 87.; Т3 Глава 26 Физиология труда – С.688 – 709. 2. Рафф Г. Секреты физиологии. Пер с англ. М. – СПб.: «Издательство БИНОМ» - « Невский диалект». 2001. – Глава 8. Физиология физической нагрузки и биоэнергетика мышечного сокращения. – С. 313 – 356. 3. Уилмор Дж.К., Костил Д.Л. Физиология спорта и двигательной активности. М.: олимпийская литература, 2001, 506с.
Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 4567; Нарушение авторского права страницы