
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Полимерное взаимодействие генов
В некоторых случаях степень фенотипического проявления зависит от количества доминантных генов, детерминирующих количественный признак. Принцип наследования некоторых признаков известен как принцип Нильсона-Эле: чем больше доминантных генов в генотипе, тем признак выраженее. Классический пример кумулятивной полимерии – наследование окраски семян пшеницы. При этом интенсивность окраски зависит от колмчества доминантных генов: в первом поколении окраска семян розовая, а во втором поколении расщепление составляет 1: 4: 6: 4: 1. Генетическая запись этого скрещивания будет выглядеть следующим образом:
А1 – наличие красного пигмента А2 – наличие красного пигмента а1 – отсутствие а2 – отсутствие
Р ♀ А1А1А2А2 ´ ♂ а1а1а2а2 G А1А2 а1а1 F1 А1а1А2а2 по генотипу: дигетерозиготные по фенотипу: розовые Результаты инбридинга Р: А1а1А2а2 ´ А1а1А2а2
Расщепление по генотипу 1: 2: 2: 1: 4: 2: 1: 2: 1 Расщепление по фенотипу 1- красные 4- тёмно розовые 6 – розовые 4- светло розовые 1- белые Таким образом, при полимерном взаимодействии наблюдается отклонение от закона независимого наследования У человека по типу количественной полимерии наследуется рост, вес, цвет кожи, артериальное давление, умственные способности. Так, если принять, что степень пигментации кожи определяется двумя парами несцепленных генов (на самом деле их гораздо больше), то, условно, людей по этому признаку можно разделить на пять фенотипов: ААВВ – негры ААВв или АаВВ – темные мулаты АаВв, ааВВ или ААвв – средние мулаты Аавв, ааВв – светлые мулаты аавв – белые Таким образом, накопление определенных аллелей в генотипе может вести к изменению выраженности признаков. Явление количественной полимерии широко распространено в природе и имеет важную роль в эволюции.
Сцепленное наследование В экспериментах Т.Моргана на дрозофиле было установлено, что развитие признаков, которые наследуются сцеплено, контролируется генами одной хромосомы. Гены окраски тела (B – серой и b – чёрной) и длины крыльев (V – нормальные и v – короткие, рудиментарные крылья) локализованы в одной паре гомологичных хромосом. Скрещивание серых мух с нормальными крыльями и чёрных мух с рудиментарными крыльями даст в первом поколении серых гибридов с нормальными крыльями:
G
F1 Серые с нормальными крыльями
При проведении анализирующего скрещивания, из F1 были выбраны самцы, т. к. известно, что у самцов дрозофил ахиазматический сперматогенез (т. е. кроссинговер не происходит и полнота сцепления генов ничем не нарушается).
P2 х
G;
F2
серые c норм. крыл черные короткокрылые 1: 1
В результате такого скрещивания на свет появляются особи двух фенотипов, аналогичные исходным родительским формам, причем в равных количествах: расщепление по фенотипу 1: 1. Рассматриваемые совместно результаты обоих скрещиваний убеждают в том, что развитие анализируемых признаков контролируется разными генами, и сцепленное наследование объясняется локализацией генов в одной хромосоме. Полнота сцепления в данном случае ничем не нарушается. Такое сцепление генов является полным.
Нарушение полного сцепления

F2; ; ;
сер. дл. чёрн. дл. сер.кор.. чёрн. кор 41, 5% 8, 5% 8, 5% 41, 5%
Процесс кроссинговера играет одну из ключевых ролей в развитии комбинативной изменчивости и является движущим фактором эволюции. В отношении отдельных идвидуумов, явление рекомбинации генов формирует уникальность фенотипа.
Таким образом, в результате кроссинговера, происходящего в профазе I первого мейотического деления образуются рекомбинантные типы гамет, процентный состав которых зависит от расстояния между неаллельными генами (в данном случае 17 морганид). Примером тесного сцепления генов у человека является наследование катаракты и полидактилии. Гены гемофилии и дальтонизма локализованы в Х- хромосоме на расстоянии 9, 8 морганид (М), т.е. подвергаются кроссинговеру, поэтому наследуются как неполностью сцепленные. Аутососмные гены резус-фактора и формы эритроцитов, расположенные друг от друга на расстоянии 3М и так же является примером неполного сцепления. Гены дефекта ногтей и коленной чашечки и групп крови сцеплены и находятся друг от друга на расстоянии 10М. Гены ночной слепоты и общей слепоты локализованы в одной хромасоме на расстоянии 50М. Наибольшее число генов удалось локализовать в X-хромосоме. Здесь их известно 95. В первой хромосоме известно 24 гена. Гены, определяющие группы крови по системе АВО, локализованы в 9-й хромосоме, определяющие группы крови по системе MN - во второй; ген Rh - фактора - в первой. В этой же хромосоме расположен ген, определяющий форму эритроцитов. В настоящее время полностью картированы 11; 17; 19; 21 и Х – хромосома человека. Составляется хромосомная карта 3-ей хромосомы. Составление хромосомных карт человека, определение локализации патологических генов в хромосомах имеет большое значение для диагностики и профилактики наследственных заболеваний человека. Результатом многолетних исследований (1911-1926) американских генетиков во главе с Т.Морганом явилась хромосомная теория наследственности. Основными ее положениями являются: 1. Каждый ген занимает в хромосоме определённое место – локус. 2. Гены в хромосоме расположены линейно в определенной последовательности. 3. Различные хромосомы содержат неодинаковое число генов. Набор генов каждой негомологичной хромосомы уникален. 4. Гены одной хромосомы образуют группу сцепления и наследуются вместе, т.е. сцеплено. 5. Число групп сцепления равно числу хромосом в гаплоидном наборе, (у дрозофилы их четыре, у кукурузы - 10, у мыши - 20, у человека: 23- группы сцепления у женщин и 24 - у мужчин). 6. Между гомологичными хромосомами может происходить обмен аллельными генами, т.е кроссинговер. 7. Частота кроссинговера прямо пропорциональна расстоянию между генами в группе сцепления. 8. За единицу расстояния между генами в группе сцепления принята особая единица - морганида (М). 1М=1% кроссинговера. Сцепленное наследование и наследование через гены цитоплазмы не вписывается в рамки законов, открытых Г. Менделем. Эти проблемы составляют особую часть фундаментальных основ классической генетики. Их изучение, возможно, не сулит сиюминутной выгоды, однако имеет большие перспективы в области генетической инженерии, картирования хромосом, позволяет решать стратегические задачи профилактики и коррекции наследственных заболеваний человека. Изучение гибридологических и цитологических аспектов сцепления генов и кроссинговера даёт неопровержимые доказательства хромосомной теории наследственности, позволяет проникнуть в тонкое строение ядра и хромосом, показывает биологический смысл кроссинговера. Сведения, полученные при изучении процессов рекомбинации открывают путь к познанию сложной структуры гена, что имеет большое значение для развития молекулярной генетики. Исследование процессов обмена генетической информацией посредством кроссинговера позволяет установить один из источников комбинативной изменчивости, представляющей важнейший фактор эволюции. Результаты теоретического и экспериментального изучения сцепленного наследования имеет большое значение для решения многих проблем генетики человека и медицинской генетики. Для человека уже определены все группы сцепления и полностью составлены генетические карты 11, 17, 19, 21 и X – хромосом. Установлена локализация многих генов, контролирующих развитие нормальных и патологических признаков. Так ген, определяющий резус – принадлежность локализован в 1-ой хромосоме, ген повышенной чувствительности к палочке Коха – в 16, гены, контролирующие синтез поверхностных эритроцитарных агглютиногенов – в 9-ой хромосоме и т.д. Хромосомная теория Т. Моргана подчёркивает факт размещения наследственного материала в хромосомах клеточного ядра. Однако, по мере развития генетики накопились данные, свидетельствующие о роли цитоплазмы в передаче наследственной информации. По материнскому типу наследуется целый ряд признаков: устойчивость к антибиотикам и другим лекарственным веществам у бактерий, пестролистность у герани, направление завитка раковины у прудовика (улитки) и даже некоторые заболевания у человека – например митохондриальные миопатии и энцефалопатии, атрофия зрительного нерва Лебера. В настоящее время разработаны критерии, позволяющие идентифицировать тип наследования через гены цитоплазмы для ряда наследственных заболеваний человека. Это имеет большое практическое значение для уточнения диагноза, назначения адекватного лечения и определение прогноза относительно потомства. Изучение ДНК-содержащих структур цитоплазмы – плазмид и эписом – имеет огромное значение для генетической инженерии, так как позволят манипулировать генами и создавать такие генотипы, которые выгодны человеку. Благодаря использованию свойств плазмид, в ДНК кишечной палочки был введён ген человеческого инсулина и налажено производство этого гормона для лечения больных сахарным диабетом. Несомненно, что ДНК органелл и плазмид цитоплазмы после тщательного изучения и картирования будет ещё более широко применяться для решения проблем медицинской генетики.
Популярное:
|
Последнее изменение этой страницы: 2016-06-04; Просмотров: 2149; Нарушение авторского права страницы



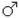
 P1 х
P1 х

 Для изучения неполного сцепления из F1 были выбраны самки (генотип). У самок во время гаметогенеза происходит кроссинговер. Поэтому дигетерозиготная особь образует дополнительные, т.е. кроссоверные сорта гамет. Вероятность их образования обусловлена вероятностью кроссинговера, т.е. зависит от расстояния между генами в группе сцепления.
Для изучения неполного сцепления из F1 были выбраны самки (генотип). У самок во время гаметогенеза происходит кроссинговер. Поэтому дигетерозиготная особь образует дополнительные, т.е. кроссоверные сорта гамет. Вероятность их образования обусловлена вероятностью кроссинговера, т.е. зависит от расстояния между генами в группе сцепления.



