
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Регенерация скелетной мышечной ткани
Ядра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами. Восстановление любых тканей организма может осуществляется за счет двух механизмов: гипертрофии и гиперплазии. Под гипертрофией подразумевают компенсаторное увеличение объема самого симпласта, в т.ч. за счет увеличения количества миофибрилл. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмолеммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Под гиперплазией понимают пролиферацию миосателлитоцитов. Сохранившиеся рядом с повреждением миосателлитоциты делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна.
40. Мышца как орган: строение, васкуляризация, эф и аф иннервация, связь мышцы с сухожилием. Скелетная мышца как орган Передача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани. Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистойсоединительной ткани — эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием. Васкуляризация. Артерии вступают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви 5—6-го порядка образуют в перимизии артериолы. В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тканевых базофилов, принимающих участие в регуляции проницаемости сосудистой стенки. Иннервация. В мышцах выявлены миелинизированные эфферентные (двигательные), афферентные (чувствительные), а также немиелинизированные вегетативные нервные волокна. Отросток нервной клетки, приносящий импульс от моторного нейрона спинного мозга, ветвится в перимизии. Каждая его ветвь проникает сквозь базальную мембрану, и у поверхности симпласта на плазмолемме образует терминали, участвуя в организации так называемой моторной бляшки, или нервно-мышечного соединения. При поступлении нервного импульса из терминали выделяется ацетилхолин — медиатор, который вызывает возбуждающий потенциал действия, распространяющееся отсюда по плазмолемме миосимпласта. Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется нервно-мышечной единицей. Характерно, что мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам. Чувствительные нервные окончания располагаются не на рабочих мышечных волокнах, а связаны со специализированными мышечными волокнами в так называемых мышечных веретенах, которые расположены в перимизии. Волокна в таких чувствительных мышечных веретенах именуются интрафузальными волокнами, а обычные рабочие мышечные волокна – экстрафузальными. Интрафузальные мышечные волокна веретен значительно тоньше рабочих. Существует два их вида — волокна с ядерной сумкой и волокна с ядерной цепочкой. Каждое мышечное волокно веретена спирально обвито терминалью чувствительного нервного волокна. В результате сокращения или расслабления рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена, соответственно изменяется тонус интрафузальных мышечных волокон. Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом.
41. Сердечная мышечная ткань. Регенерация. Сердечная мышечная ткань Гистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани — симметричные участки висцерального листка спланхнотома в шейной части зародыша — так называемые миоэпикардиалъные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 3 вида кардиомиоцитов: 1. рабочие, или типичные, или же сократительные, кардиомиоциты, 2. атипичные кардиомиоциты (сюда входят пейсмекерные, проводящие и переходные кардиомиоциты, а также 3. секреторные кардиомиоциты. Рабочие ( сократительные ) кардиомиоциты образуют свои цепочки. Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают гормон - натрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Сократительные кардиомиоциты имеют удлиненную (100—150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски. Кардиомиоциты могут ветвиться и образуют трехмерную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Миофибриллы слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены, контактируют с мембранами гладкой эндоплазматической (т.е. саркоплазматической) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта. Кардиомиоциты соединяются друг с другом своими торцевыми концами. Здесь образуются так называемые вставочные диски: эти участки выглядят как тонкие пластинки при увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (или щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений. Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани.
42. Хар-ка нервной ткани. Источники развития. Нейроциты: функции, строение, морфолог и функциональн классификация. Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и его передачи. Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой. В нервной ткани выделяют два типа клеток – нервные и глиальные. Нервные клетки (нейроны, или нейроциты) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию. Нейроглия обеспечивает существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции. Развитие
Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней кожной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки не входит в состав ни нервной трубки, ни кожной эктодермы, а образует скопления по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся между нервной трубкой и кожной эктодермой, — это нервный гребень (или ганглиозная пластинка). Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных и автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и некоторым видам глии: нейролеммоцитам (шванновским клеткам), клеткам-сателлитам ганглиев. Из нервного гребня развиваются также клетки мозгового вещества надпочечников, меланоциты кожи, часть клеток APUD-системы, сенсорные клетки каротидных телец. В формировании ганглиев V, VII, IX и X пар черепных нервов принимают участие, кроме нервного гребня, также нейрогенные плакоды, представляющие собой утолщения эктодермы по бокам формирующейся нервной трубки в краниальном отделе зародыша. Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных, или нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: · внутренняя - вентрикулярная (или эпендимная) зона, · вокруг нее – субвентрикулярная зона, · затем промежуточная (или плащевая, или же мантийная, зона) и, наконец, · наружная - краевая (или маргинальная) зона нервной трубки. Вентрикулярная (эпендимная), внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии. Субвентрикулярная зона состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая, или мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон — нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. Способность к делению не утрачивают полностью и зрелые глиоциты. Новообразование нейронов прекращается в раннем постнатальном периоде. Поскольку число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в среднем в течение всего пренатального периода в 1 мин формируется 2, 5 миллиона нейронов. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга. Маргинальная зона (или краевая вуаль) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых областях головного мозга клетки плащевого слоя мигрируют дальше, образуя кортикальные пластинки — скопления клеток, из которых формируется кора большого мозга и мозжечка (т.е. серое вещество). По мере дифференцировки нейробласта, изменяется субмикроскопическое строение его ядра и цитоплазмы. Специфическим признаком начавшейся специализации нервных клеток следует считать появление в их цитоплазме тонких фибрилл — пучков нейрофиламентов и микротрубочек. Количество нейрофиламентов, содержащих белок — нейрофиламентный триплет, в процессе специализации увеличивается. Тело нейробласта постепенно приобретает грушевидную форму, а от его заостренного конца начинает развиваться отросток — аксон. Позднее дифференцируются другие отростки — дендриты. Нейробласты превращаются в зрелые нервные клетки — нейроны. Между нейронами устанавливаются контакты (синапсы). В процессе дифференцировки нейронов из нейробластов различают до-медиаторный и медиаторный периоды. Для домедиаторного периода характерно постепенное развитие в теле нейробласта органелл синтеза — свободных рибосом, а затем эндоплазматической сети. В медиаторном периоде у юных нейронов появляются первые пузырьки, содержащие нейромедиатор, а в дифференцирующихся и зрелых нейронах отмечаются: значительное развитие органелл синтеза и секреции, накопление медиаторов и поступление их в аксон, образование синапсов. Несмотря на то, что формирование нервной системы завершается только в первые годы после рождения, известная пластичность центральной нервной системы сохраняется до старости. Эта пластичность может выражаться в появлении новых терминалей и новых синаптических связей. Нейроны центральной нервной системы млекопитающих способны формировать новые ветви и новые синапсы. Пластичность проявляется в наибольшей степени в первые годы после рождения, но частично сохраняется и у взрослых — при изменении уровней гормонов, обучении новым навыкам, травме и других воздействиях. Хотя нейроны постоянны, их синаптические связи могут модифицироваться в течение всей жизни, что может выражаться, в частности, в увеличении или уменьшении их числа. Пластичность при малых повреждениях мозга проявляется в частичном восстановлении функций. В популяции нейронов, начиная с ранних стадий развития нервной системы и в течение всего онтогенеза, имеет место массовая гибель клеток. Эта запрограммированная физиологическая гибель клеток наблюдается как в центральной, так и в периферической нервной системе. У человека ежегодно погибает около 10 млн нервных клеток. Нейроны, или нейроциты, — специализированные клетки нервной системы, ответственные за получение, обработку и передачу сигнала (на: другие нейроны, мышечные или секреторные клетки). Нейрон является морфологически и функционально самостоятельной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими нейронами, образуя рефлекторные дуги — звенья цепи, из которой построена нервная система. В зависимости от функции в рефлекторной дуге различают три типа нейронов: · афферентные · ассоциативные · эфферентные Афферентные (или рецепторные, чувствительные) нейроны воспринимают импульс, эфферентные (или двигательные) передают его на ткани рабочих органов, побуждая их к действию, а ассоциативные (или вставочные) осуществляют связь между нейронами. Подавляющее большинство нейронов (99, 9%) - ассоциативные. Нейроны отличаются большим разнообразием форм и размеров. Например, диаметр тел клеток-зерен коры мозжечка 4—6 мкм, а гигантских пирамидных нейронов двигательной зоныкоры большого мозга — 130—150 мкм. Нейроны состоят из тела (или перикариона) и отростков: одного аксона и различного числа ветвящихся дендритов. По количеству отростков различают три типа нейронов: · · мультиполярные (большинство) и · униполярные нейроны. Униполярные нейроны имеют только аксон (у высших животных и человека обычно не встречаются). Биполярные - имеют аксон и один дендрит. Мультиполярные нейроны (подавляющее большинство нейронов) имеют один аксон и много дендритов. Разновидностью биполярных нейронов является псевдо-униполярный нейрон, от тела которого отходит один общий вырост — отросток, разделяющийся затем на дендрит и аксон. Псевдоуниполярные нейроны присутствуют в спинальных ганглиях, биполярные — в органах чувств. Большинство нейронов - мультиполярные. Их формы чрезвычайно разнообразны. Аксон и его коллатерали оканчиваются, разветвляясь на несколько веточек, называемых телодендронами, последние заканчиваются терминальными утолщениями. Трехмерная область, в которой ветвятся дендриты одного нейрона, называется дендритным полем нейрона. Дендриты представляют собой истинные выпячивания тела клетки. Они содержат те же органеллы, что и тело клетки: глыбки хроматофильной субстанции (т.е. гранулярной эндоплазматической сети и полисом), митохондрии, большое количество нейротубул (или микротрубочек) и нейрофиламентов. За счет дендритов рецепторная поверхность нейрона увеличивается в 1000 и более раз. Аксон — это отросток, по которому импульс передается от тела клетки. Он содержит митохондрии, нейротубулы и нейрофиламенты, а также гладкую эндоплазматическую сеть. Подавляющее большинство нейронов человека содержит одно округлое светлое ядро, расположенное в центре клетки. Двуядерные и тем более многоядерные нейроны встречаются крайне редко. Плазмолемма нейрона является возбудимой мембраной, т.е. обладает способностью генерировать и проводить импульс. Ее интегральными белками являются белки, функционирующие как ионно-избирательные каналы, и рецепторные белки, вызывающие реакции нейронов на специфические стимулы. В нейроне мембранный потенциал покоя равен —60 —70 мВ. Потенциал покоя создается за счет выведения Na+ из клетки. Большинство Na+- и К+-каналов при этом закрыты. Переход каналов из закрытого состояния в открытое регулируется мембранным потенциалом. В результате поступления возбуждающего импульса на плазмолемме клетки происходит частичная деполяризация. Когда она достигает критического (порогового) уровня, натриевые каналы открываются, позволяя ионам Na+ войти в клетку. Деполяризация усиливается, и при этом открывается еще больше натриевых каналов. Калиевые каналы также открываются, но медленнее и на более продолжительный срок, что позволяет К+ выйти из клетки и восстановить потенциал до прежнего уровня. Через 1—2 мс (т.н. рефрактерный период) каналы возвращаются в нормальное состояние, и мембрана может вновь отвечать на стимулы. Итак, распространение потенциала действия обусловлено вхождением в нейрон ионов Na+, которые могут деполяризовать соседний участок плазмолеммы, что в свою очередь создает потенциал действия на новом месте. При окрашивании нервной ткани анилиновыми красителями в цитоплазме нейронов выявляется хроматофильная субстанция в виде базофильных глыбок и зерен различных размеров и форм (другие названия хроматофильной субстанции - тигроид, тельца Ниссля). Базофильные глыбки локализуются в перикарионах и дендритах нейронов, но никогда не обнаруживаются в аксонах и их конусовидных основаниях — аксональных холмиках. Базофилия глыбок объясняется высоким содержанием рибонуклеопротеидов. Каждая глыбка хроматофильной субстанции состоит из цистерн гранулярной эндоплазматической сети, свободных рибосом и полисом. Для поддержания целостности нейронов и выполнения ими функций нейронам требуется огромное количество белков. Для аксонов, не имеющих органелл белкового синтеза, характерен постоянный ток цитоплазмы от перикариона к терминалям со скоростью 1—3 мм в сутки. Возрастные изменения нейронов сопровождаются накоплением липофусцина, разрушением крист митохондрий. Липофусцин — «пигмент старения» — желто-бурого цвета липопротеидной природы, представляющий собой остаточные тельца (т.е. телолизосомы) с продуктами непереваренных структур.
43. Нервные волокна: определение, строение и функции миели и безмиелин волокон. Регенерация. Нервные волокна Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают миелиновые и безмиелиновые нервные волокна. Отросток нервной клетки в нервном волокне называют осевым цилиндром, или аксоном, так как чаще всего (за исключением чувствительных нервов) в составе нервных волокон находятся именно аксоны. В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической — нейролеммоцитами Шванна. Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон, на которой как бы подвешен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, или перехваты Ранвье. В процессе миелинизации аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается (как бы накручивается) на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой. Осевой цилиндр в этом месте частично прикрыт интердигитирующими отростками нейролеммоцитов. Оболочка аксона ( аксолемма ) обладает в области перехвата значительной электронной плотностью. Отрезок волокна между смежными перехватами называется межузловым сегментом. Длина межузлового сегмента, так же как и толщина миелинового слоя, зависит от толщины осевого цилиндра. Насечка миелина (Шмидта—Лантермана) представляет собой участок миелинового слоя, где завитки мезаксона лежат неплотно друг к другу, образуя спиральный туннель, идущий снаружи внутрь и заполненный цитоплазмой нейролеммоцита, т.е. место расслоения миелина. Снаружи от нейролеммоцита располагается базальная мембрана. Миелиновые волокна центральной нервной системы не имеют насечек миелина, а нервные волокна не окружены базальными мембранами. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1—2 м/с, тогда как толстые миелиновые — со скоростью 5—120 м/с. В безмиелиновом волокне волна деполяризации мембраны идет по всей аксолемме, не прерываясь, а в миелиновом возникает только в области перехватов. Таким образом, для миелиновых волокон характерно сальтаторное проведение возбуждения, т.е. прыжками. Между перехватами идет электрический ток, скорость которого выше, чем прохождение волны деполяризации по аксолемме. Регенерация зависит от места травмы. Как в центральной, так и в периферической нервной системе погибшие нейроны не восстанавливаются. Полноценной регенерации нервных волокон в центральной нервной системе обычно не происходит, но нервные волокна в составе периферических нервов обычно хорошо регенерируют. При этом нейролеммоциты периферического отрезка и ближайшего к области травмы участка центрального отрезка пролиферируют и выстраиваются компактными тяжами. Осевые цилиндры центрального отрезка дают многочисленные коллатерали, которые растут со скоростью 1—3 мм в сутки вдоль нейролеммальных тяжей, создавая, таким образом, избыточный рост нервных волокон. Выживают только те волокна, которые достигают соответствующих окончаний. Остальные дегенерируют. Если существует препятствие для врастания аксонов центрального отрезка нерва в тяжи нейролеммоцитов периферического отрезка (например, при наличии рубца), аксоны центрального отрезка растут беспорядочно и могут образовать клубок, называемый ампутационной невромой. Поврежденные нервные волокна головного и спинного мозга не регенерируют. Возможно, регенерации нервных волокон в центральной нервной системе не происходит потому, что глиоциты без базальной мембраны лишены хемотаксических факторов, необходимых для проведения регенерирующих аксонов. Однако при малых травмах центральной нервной системы возможно частичное восстановление ее функций, обусловленное пластичностью нервной ткани. 44. Нейроглия: классификация, строение и тдтд Нейроглия Нейроны — это высокоспециализированные клетки, существующие и функционирующие в строго определенной среде. Такую среду им обеспечивает нейроглия. Нейроглия выполняет следующие функции: опорную, трофическую, разграничительную, поддержание постоянства среды вокруг нейронов, защитную, секреторную. Различают глию центральной и периферической нервной системы. Клетки глии центральной нервной системы делятся на макроглию и микроглию. Макроглия Макроглия развивается из глиобластов нервной трубки и включает: эпендимоциты, астроциты и олигодендроглиоциты. Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Эти клетки цилиндрической формы. Они образуют слой типа эпителия, носящий название эпендимы. Между соседними клетками эпендимы имеются щелевидные соединения и пояски сцепления, но плотные соединения отсутствуют, так что цереброспинальная жидкость может проникать между эпендимоцитами в нервную ткань. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости. Базальная поверхность большинства эпендимоцитов ровная, но некоторые клетки имеют длинный отросток, идущий глубоко в нервную ткань. Такие клетки называются таницитами. Они многочисленны в дне III желудочка. Считается, что эти клетки передают информацию о составе цереброспинальной жидкости на первичную капиллярную сеть воротной системы гипофиза. Эпендимный эпителий сосудистых сплетений желудочков продуцирует цереброспинальную жидкость (ликвор). Астроциты — клетки отростчатой формы, бедные органеллами. Они выполняют в основном опорную и трофическую функции. Различают два типа астроцитов - протоплазматические и волокнистые. Протоплазматические астроциты локализуются в сером веществе центральной нервной системы, а волокнистые астроциты - преимущественно в белом веществе. Протоплазматические астроциты характеризуются короткими сильно ветвящимися отростками и светлым сферическим ядром. Отростки астроцитов тянутся к базальным мембранам капилляров, к телам и дендритам нейронов, окружая синапсы и отделяя (изолируя) их друг от друга, а также к мягкой мозговой оболочке, образуя пиоглиальную мембрану, граничащую с субарахноидальным пространством. Подходя к капиллярам, их отростки образуют расширенные «ножки», полностью окружающие сосуд. Астроциты накапливают и передают вещества от капилляров к нейронам, захватывают избыток экстрацеллюлярного калия и других веществ, таких как нейромедиаторы, из экстрацеллюлярного пространства после интенсивной нейрональной активности. Олигодендроциты – имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах, причем, в противоположность аналогичным клеткам периферической нервной системы – нейролеммоцитам, один олигодендроглиоцит может участвовать в миелинизации сразу нескольких аксонов. Микроглия Микроглия представляет собой фагоцитирующие клетки, относящиеся к системе мононуклеарных фагоцитов и происходящие из стволовой кроветворной клетки (возможно, из премоноцитов красного костного мозга). Функция микроглии — защита от инфекции и повреждения, и удаление продуктов разрушения нервной ткани. Клетки микроглии характеризуются небольшими размерами, телами продолговатой формы. Их короткие отростки имеют на своей поверхности вторичные и третичные ответвления, что придает клеткам «колючий» вид. Описанная морфология характерна для типичной (ветвистой, или покоящейся) микроглии полностью сформированной центральной нервной системы. Она обладает слабой фагоцитарной активностью. Ветвистая микроглия встречается как в сером, так и в белом веществе центральной нервной системы. В развивающемся мозгу млекопитающих обнаруживается временная форма микроглии — амебоидная микроглия. Клетки амебоидной микроглии формируют выросты – филоподии и складки плазмолеммы. В их цитоплазме присутствуют многочисленные фаголизосомы и пластинчатые тельца. Амебоидные микроглиальные тельца отличаются высокой активностью лизосомальных ферментов. Активно фагоцитирующая амебоидная микроглия необходима в раннем постнатальном периоде, когда гематоэнцефалический барьер еще не вполне развит и вещества из крови легко попадают в центральную нервную систему. Считают также, что она способствует удалению обломков клеток, появляющихся в результате запрограммированной гибели избыточных нейронов и их отростков в процессе дифференцировки нервной системы. Полагают, что, созревая, амебоидные микроглиальные клетки превращаются в ветвистую микроглию. Реактивная микроглия появляется после травмы в любой области мозга. Она не имеет ветвящихся отростков, как покоящаяся микроглия, не имеет псевдоподий и филоподий, как амебоидная микроглия. В цитоплазме клеток реактивной микроглии присутствуют плотные тельца, липидные включения, лизосомы. Есть данные о том, что реактивная микроглия формируется вследствие активации покоящейся микроглии при травмах центральной нервной системы. Рассмотренные выше глиальные элементы относились к центральной нервной системе. Глия периферической нервной системы в отличие от макроглии центральной нервной системы происходит из нервного гребня. К периферической нейроглии относятся: нейролеммоциты (или шванновские клетки) и глиоциты ганглиев (или мантийные глиоциты). Нейролеммоциты Шванна формируют оболочки отростков нервных клеток в нервных волокнах периферической нервной системы. Мантийные глиоциты ганглиев окружают тела нейронов в нервных узлах и участвуют в обмене веществ этих нейронов.
45. Нервные окончания: классифик, строение и тд Нервные окончания Нервные волокна заканчиваются концевыми аппаратами — нервными окончаниями. Различают три группы нервных окончаний: · межнейрональные синапсы, осуществляющие связь нейронов между собой; · эффекторные окончания (эффекторы), передающие нервный импульс на ткани рабочего органа (на мышечные или железистые клетки) · рецепторные (или аффекторные, или же чувствительные) окончания Межнейрональные синапсы
Популярное:
|
Последнее изменение этой страницы: 2016-05-30; Просмотров: 1123; Нарушение авторского права страницы
 Нейруляция (схема).
Нейруляция (схема). биполярные,
биполярные, 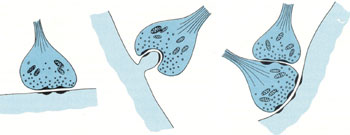 Синапсы — это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. Синапсы определяют направление проведения импульса. Если раздражать аксон электрическим током, импульс пойдет в обоих направлениях; но импульс, идущий в сторону тела нейрона и его дендритов, не может быть передан на другие нейроны. Только импульс, достигающий терминалей аксона, с помощью синапсов может передать возбуждение на другой нейрон, мышечную или железистую клетку. В зависимости от способа передачи импульса синапсы могут быть химическими или электрическими (электротоническими).
Синапсы — это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. Синапсы определяют направление проведения импульса. Если раздражать аксон электрическим током, импульс пойдет в обоих направлениях; но импульс, идущий в сторону тела нейрона и его дендритов, не может быть передан на другие нейроны. Только импульс, достигающий терминалей аксона, с помощью синапсов может передать возбуждение на другой нейрон, мышечную или железистую клетку. В зависимости от способа передачи импульса синапсы могут быть химическими или электрическими (электротоническими).