|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Формы и размеры бактериальных организмов и их краткая характеристикаСтр 1 из 5Следующая ⇒
ВВЕДЕНИЕ Изучение морфолого-анатомических признаков бактерий помогает установить систематическое положение той или иной группы бактерий и, в конечном итоге определить их практическую значимость. Небольшие размеры бактерий долгое время казались несовместимыми с какой-либо клеточной дифференцировкой. Использование новейших методов исследования позволило выделить клеточные структуры изучить их физиологические функции. Большую роль в изучении бактерий сыграли косвенные методы, основанные на способности клетки реагировать на иммунные антитела. Оказалось, что реагируют в клетке белки с определенной формой и расположением полипептидной цепи. Это позволило изучить то, что лежит за пределами видимости микроскопа. Исследования показали, что тело микробной клетки морфологически дифференцировано. Снаружи оно покрыто оболочкой или правильнее сказать клеточной стенкой, которая предохраняет клетку, придает ей определенную форму, является барьером для белков и полисахаридов, обладает определенной биохимической активностью. Пример: в оболочке клеток туберкулеза обнаружен фактор, тормозящий продвижение лейкоцитов к очагу и придает упругость клетке, но допускает свободное движение связанное с изгибами тела. Один из основных признаков прокариотической клетки — отсутствие внутреннего разделения, обеспечиваемого элементарны-ми мембранами. По сути, единственная мембранная система прокариотической клетки — ЦПМ, входящая в состав клеточной оболочки, часто сложно устроенная и проникающая поперечными складками глубоко в цитоплазму. В составе любой стабильной бактериальной клетки выделяют условные компартменты — поверхностные структуры, клеточную оболочку и цитоплазму.
ЛИТЕРАТУРНЫЙ ОБЗОР Цитоплазматическая мембрана
Два слоя фосфолипидных молекул, обращенных гидрофобными полюсами друг к другу и покрытых двумя слоями молекул глобулярного белка. С ЦПМ связаны многие ферменты и белки, участвующие в переносе питательных веществ, а также ферменты и переносчики электронов конечных стадий биологического окисления (дегидрогеназы, цитохром-ная система, АТФ-аза). На ЦМП локализуются ферменты, катализирующие синтез пептидогликана, белков клеточной стенки, собственных структур. Мембрана является также местом превращения энергии при фотосинтезе. Периплазматическое пространство Периплазматическое пространство (периплазма) представляет собой зону между клеточной стенкой и ЦПМ. Толщина периплазмы составляет около 10 нм, объем зависит от условий среды и прежде всего от осмотических свойств раствора. Периплазма может включать до 20 % всей находящейся в клетке воды, в ней локализуются некоторые ферменты (фосфатазы, пермеазы, нуклеазы и др.) и транспортные белки -переносчики соответствующих субстратов. Цитоплазма Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий. Та часть цитоплазмы, которая имеет гомогенную коллоидную консистенцию и содержит растворимые РНК, ферменты, субстраты и продукты обмена веществ, обозначается как цитозоль. Другая часть цитоплазмы представлена различными структурными элементами: мезосомами, рибосомами, включениями, нуклеоидом, плазмидами. Рибосомы - субмикроскопические рибонуклеопротеиновые гранулы диаметром 15-20 нм. В рибосомах находится примерно 80-85 % всей бактериальной РНК. Рибосомы прокариот имеют константу седиментации 70 S. Они построены из двух частиц: 30 S (малая субъединица) и 50 S (большая субъединица)(рис.1.6).
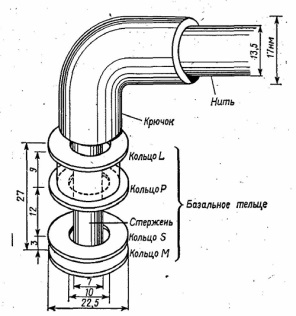
Рис. 1.6. Рибосома (а) и ее субчастицы - большая (б) и малая (в) Рибосомы служат местом синтеза белка. Цитоплазматические включения Нередко в цитоплазме бактерий обнаруживаются различные включения, которые образуются в процессе жизнедеятельности: капельки нейтральных липидов, воска, серы, гранулы гликогена, в-гидроксимасляной кислоты (особенно у рода Bacillus). Гликоген и в-гидроксимасляная кислота служат для бактерий запасным источником энергии. У некоторых бактерий в цитоплазме находятся кристаллы белковой природы, обладающие ядовитым действием на насекомых. Некоторые бактерии способны накапливать фосфорную кислоту в виде гранул полифосфата (зерна волютина, метахроматические зерна). Они играют роль фосфатных депо и выявляются в виде плотных образований в форме шара или эллипса, располагающихся в основном у полюсов клетки. Обычно на полюсах бывает по одной грануле. Нуклеоид Нуклеоид - ядерный аппарат бактерий. Представлен молекулой ДНК, соответствующей одной хромосоме. Она замкнута, располагается в ядерной вакуоле, не имеет ограничивающей от цитоплазмы мембраны. С ДНК связано небольшое количество РНК и РНК-полимеразы. ДНК свернута вокруг центрального стержня, состоящего из РНК, и представляет собой высокоупорядоченную компактную структуру. Хромосомы большинства прокариот имеют молекулярную массу в пределах 1-3 х109, константу седиментации 1300-2000 S. Молекула ДНК включает 1, 6х10 нуклеотидных пар. Различия в генетическом аппарате прокариотических и эукариотических клеток обусловливают его название: у первых - нуклеоид (образование, подобное ядру), в отличие от ядра у вторых. В нуклеоиде бактерий содержится основная наследственная информация, которая реализуется в синтезе специфических белковых молекул. С ДНК бактериальной клетки связаны системы репликации, репарации, транскрипции и трансляции. Нуклеоид в прокариотической клетке может быть выявлен в окрашенных препаратах с помощью светового или фазово-контрастного микроскопа.э У многих бактерий в цитоплазме обнаружены внехромосомные генетические элементы - плазмиды. Они представляют собой замкнутые в кольца двухцепочечные ДНК, состоящие из 1500-40000 пар нуклеотидов и содержащие до 100 генов. Капсула Капсула - слизистый слой клеточной стенки бактерий, состоящий из полисахаридов или полипептидов. Микрокапсулу (толщиной менее 0, 2 мкм) способны формировать большинство бактерий. Жгутики Жгутики выполняют роль органа движения, позволяющего бактериям передвигаться со скоростью 20-60 мкм/сек. Бактерии могут иметь один или несколько жгутиков, располагающихся по всей поверхности тела либо собранных в пучки у одного полюса, у разных полюсов. Толщина жгутиков в среднем составляет 10-30 нм, а длина достигает 10-20 мкм. Основу жгутика составляет длинная спиральная нить (фибрилла), которая у поверхности клеточной стенки переходит в утолщенную изогнутую структуру - крюк и прикрепляется к базальной грануле, вмонтированной в клеточную стенку и ЦПМ (рис. 1.7).
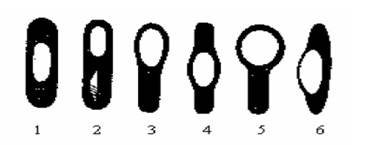
Удаление пептидогликанового слоя клеточной стенки ведет к потере способности бактерий к движению, хотя жгутики при этом остаются неповрежденными. Жгутики почти полностью состоят из белка флагеллина с некоторым содержанием углеводов и РНК. Споры Некоторые бактерии в конце периода активного роста способны образовывать споры. Этому предшествует обеднение среды питательными веществами, изменение ее рН, накопление ядовитых продуктов метаболизма. Как правило, одна бактериальная клетка образует одну спору - локализация спор различна (центральная, терминальная, субтерминальная - рис. 1.8).
Рис. 1.8. Типичные формы спорообразующих клеток. Если размеры спор не превышают поперечного размера палочковидной бактерии, то последняя называется бациллой. Когда диаметр споры больше - бактерии имеют форму веретена и носят название клостридий. По химическому составу различие спор от вегетативных клеток состоит лишь в количественном содержании химических соединений. Споры содержат меньше воды и больше липидов. В состоянии споры микроорганизмы метаболически неактивны, выдерживают высокую температуру (140-150°С) и воздействие химических дезинфицирующих веществ, длительно сохраняются в окружающей среде. Попадая в питательную среду, споры прорастают в вегетативные клетки. Процесс прорастания спор включает три стадии: активации, начальной стадии и стадии роста. К активирующим агентам, нарушающим состояние покоя, относят повышенную температуру, кислую реакцию среды, механические повреждения и др. Спора начинает поглощать воду и с помощью гидролитических ферментов разрушает многие собственные структурные компоненты. После разрушения наружных слоев наступает период формирования вегетативной клетки с активацией биосинтеза, заканчивающейся делением клетки.
2.1 Спорообразование и его биологическая роль Движение бактерий не является особенностью всех бактерий, только некоторые способны двигаться. Органом движения является жгутик. Подвижные спирохеты движутся за счет ритмических изгибов всего тела. Для миксобактерий характерно реактивное движение за счет выделяющейся слизи. Характер движения у жгутиковых организмов зависит от типа жгутикования. Бактерии с полярным жгутиком всегда движутся по прямой линии, только изредка совершая легкие колебательные движения. Движение перитрихов беспорядочно и связано с оживленным кувырканием. У лофотрихов жгутики перемещаются подобно корабельному винту. Скорость движения у различных бактерий различна. Некоторые виды при благоприятных условиях могут двигаться на расстояние в 10 раз превышающее размеры клетки. Процесс спорообразования у бактерий является стадией развития. Спора играет роль защитного приспособления, которым природа снабдила их для борьбы с неблагоприятными условиями. Способностью образовывать споры - особые тельца эллиптической и округлой формы обладают палочковидные бактерии. На образование споры затрачивается или все содержимое клетки или большая часть протоплазматического содержимого. Спора образуется в результате уплотнения протоплазмы в определенном участке, содержащим ядерную субстанцию. Эта часть названа А.А. Имшеницким спорогенной зоной. В дальнейшем из этого участка вначале образуется проспора, которая далее покрывается специальными оболочками и превращается в спору. Длительность процесса 1, 5-24 часа. В начале фазы спора хорошо окрашивается анилиновыми красками, а по мере созревания она становится более плотной и трудноокрашиваемой, что (связано) объясняется малопроницаемой наружной оболочкой. Однако, такое объяснение не вполне удовлетворительно, т.к. в зрелом состоянии оболочка споры окрашивается, а содержимое остается не окрашенным. Это дает основание предположить, что цитоплазма изменила физико-химические свойства. А именно: вода находится в связанном состоянии, благодаря чему спора приобретает теплоустойчивость, увеличивается содержание Са и Mg, что тоже повышает стойкость к нагреванию. В дальнейшем оболочка споры раздваивается: внутреннее - интина - участвует в прорастании споры и на ее базе строится оболочка новой вегетативной клетки. Наружная оболочка -экзина выполняет защитную функцию. По мере созревания споры, вегетативная клетка отмирает. Малая активность ферментов, плохая проницаемость наружной оболочки ограничивают обмен спор с внешней средой, что позволяет им сохранить жизнеспособность десятки и сотни лет. Жизнеспособные споры были выделены из египетских мумий, трупов мамонтов. Обычно к спорообразованию бациллы приступают при истощении питательной среды или при накоплении в среде продуктов обмена. Механизм спорообразования не ясен. В благоприятных условиях спора прорастает. Сначала она набухает, увеличивается ее размеры, происходит активизация ферментов, разрывается нарушенная оболочка и проросток выходит наружу. Переход из споры в вегетативную клетку может происходить за 40-50 мин. Обычно споры прорастают полярно, на одном или на обоих концах, иногда экваториально поперечно продольной оси, реже наблюдается косое прорастание. Некоторые бациллы при определенных условиях теряют способность образовывать споры (например: при накоплении ядовитых веществ в среде возникают целые поколения - аспорогенные расы). Таким образом, спора - защитное приспособление не имеющее никакого отношения к размножению. Размножение бактерий Основным путем размножения у бактерий является бесполое размножение. Однако у некоторых бактерий встречается и половое размножение - конъюгация. Бесполое размножение: молодая клетка растет, достигает определенной величины и перпендикулярно ее длине образуется внутренняя перегородка. Затем перегородка расщепляется надвое и получается две дочерние клетки (фактически образуется две клетки: материнская и дочерняя). Скорость размножения зависит от условий, при благоприятных внешних факторах деление отмечается через каждые 20-30 мин, а у термофилов даже через 5 минут. Разновидностью бесполого размножения является почкование, которое имеет место наряду с делением у микобактерий. Скорость размножения настолько велика, что теоретически микробные клетки заполнили бы все моря и океаны за несколько суток. Однако различные факторы тормозят процесс размножения бактерий: истощение питательных веществ в среде, накопление продуктов обмена, поедание их простейшими, растворение фагами, действие физических и химических факторов. Некоторые бактерии имеет в своем развитии ряд последовательных изменений, которые можно назвать циклом развития. Пример: молодая сенная палочка (bac.subtilis) несет много жгутиков, которые она перед деление сбрасывает и усиленно делится, образуя длинные цепочки палочек. Затем цепочки распадаются на отдельные клетки и каждая из них вновь размножается. После многократного размножения клетки переходят к спорообразованию. При благоприятных условиях спора прорастает и образуется подвижная палочка. Половое размножение - конъюгация впервые открыто Ледербергом и Татум (1946) и доказано генетическим методом. После их открытия было подтверждено с помощью электрического микроскопа. Суть коньюгации: клетка - донор, реципиент сближаются, и между ними образуется цитоплазматический мостик, по которому генетический материал донора переходит в клетку реципиент. После расхождения эти клетки могут делиться бесполым путем. Биологический смысл этого процесса в том, что с передачей генетического материала передаются и определенные свойства, факторы, которыми не обладала клетка реципиент. Так могут чередоваться «факторы устойчивости» к стрептомицину, тетрациклину и др. веществам, в результате бактерии приобретают устойчивость к антибиотикам и другим терапевтическим средствам. Физиология бактерий Жизнь бактерий, как и других живых существ, в упрощённом виде сводится к таким последовательным событиям: собственное воспроизведение -» обеспечение жизненных функций -» воспроизведение потомства. Нормальная реализация этого круговорота жизни для любого вида бактерий возможна лишь при развитии адекватных ответных реакций на любые воздействия окружающей среды, что во многом обеспечивается генетической информацией организма. Жизнь бактерий, равно как и всех живых существ, характеризуется ростом и размножением, то есть увеличением живой массы отдельной особи и популяции в целом за счет ассимиляции веществ, находящихся вне клетки. В данном аспекте нет никакой существенной разницы между питанием паразитической бактерии или автотрофной бактерии, использующей, подобно высшим растениям, углекислоту и минеральные вещества. Во всех случаях процесс питания сводится к усвоению экзогенных субстратов и превращению их (через каскад сложных реакций) в вещества, необходимые для нормальной жизнедеятельности бактерий. Питание бактерий Пишей принято называть любое вещество, которое, попав в организм, служит источником энергии или пластическим материалом для синтеза молекул, используемых для нужд организма. Большинство животных, включая человека, способно заглатывать и переваривать плотные частички пищи в основном за счёт их гидролиза. Подобный тип питания известен как голо- зойныЁ, а организмы — голозои [от греч. hobs, полноценный, + zoikos, относящийся к животным]. Бактерии не способны захватывать твердофазные объекты, поэтому утилизируют питательные вещества в виде относительно простых молекул из водных растворов. Подобный тип питания, присущий также всем растениям, известен как голофитный, то есть бактерии — голофиты [от греч. holos, полноценный, + phytikos, относящийся к растениям]. Тем не менее многие бактерии способны утилизировать твёрдую пищу с помощью так называемого внешнего питания, реализуемого вне клеток, то есть бактериям также присущ и голозойный тип питания. Для этого они имеют мощный ферментативный потенциал, хотя иногда секретируемые ферменты могут полностью инактивироваться в результате разведения, под действием конвекционных токов и других факторов. Контакт пищеварительных ферментов с экзогенным субстратом приводит к образованию низкомолекулярных продуктов, проникающих через клеточную стенку в цитоплазму. Начиная с этого момента, процессы их усвоения (метаболизма) в растительных и животных клетках протекают удивительно сходно. Клетки всех живых существ, от самых примитивных форм до высокоразвитых животник и растений, не только состоят из одних и тех же веществ, но и используют одни и те же механизмы для получения энергии и для роста. По сравнению с этим фундаментальным биохимическим единством существующие различия и отклонения кажутся незначительными. Вода Бода составляет около 80% массы бактерий. Рост и развитие бактерий облигатно зависят от наличия воды, так как все химические реакции, протекающие в живых организмах, реализуются в водной среде. Для нормального роста и развития микроорганизмов необходимо присутствие воды в окружающей среде. Для бактерий содержание воды в субстрате должно быть более 20%. Вода должна находиться в доступной форме: в жидкой фазе в интервале температур от 2 до 60 °С; этот интервал известен как биокипетическая зона. Хотя в химическом отношении вода весьма устойчива, продукты её ионизации — ионы Н+ и ОН” оказывают очень большое влияние на свойства практически всех компонентов клетки (белков, нуклеиновых кислот, липидов и т.д.). Так, каталитическая активность ферментов в значительной мере зависит от концентрации ионов Н+ и ОН". Усваиваемые соединения Основные соединения, усваиваемые бактериальной клеткой, — углеводы, аминокислоты, органические кислоты, жирные кислоты, минеральные вещества, витамины и др. Бактериям совершенно безразличны источники питательных веществ; образно говоря, они «лишены вкуса и не страдают несварением желудка». Более того, бактерии иногда утилизируют вещества, не пригодные для животных клеток (например, карболовую кислоту, парафин, мыло и др.). Подобно прочим формам жизни, бактерии нуждаются в одних и тех же макроэлементах — С, Н, О, N, Р, S, К, Са, Mg, Fe. Микроэлементы (следовые элементы) — Mn, Mo, Zn, Си, Со, Ni, Va, В, С], Na, Se, Si, Wo — не нужны каждому организму, но бактериям они необходимы для синтеза коферментов либо поддержания специфического типа метаболизма. Например, для оптимального роста некоторые бактерии нуждаются в высоких концентрациях Na+; их называют галофилами [от греч. hals, соль]. Помимо источников углерода, энергии и элементов минерального питания, многие микроорганизмы нуждаются в некоторых дополнительных веществах, называемых факторами роста. Количественная потребность в питательных элементах и их содержание у различных бактерий варьируют, но принципиально химический состав бактериальной клетки сходен с другими живыми клетками (исключением является отсутствие у бактерий стеролов). Пути поступления веществ в бактериальную клетку Для того чтобы питательные вещества могли подвергнуться соответствующим превращениям в клетке, они прежде всего должны в неё проникнуть. Но большинство бактерий обитает в условиях, мало пригодных для поддержания строгих соотношений воды, неорганических и органических веществ, без которых их жизнь просто невозможна. Клеточная стенка бактерий не является существенным барьером для небольших молекул и ионов, но задерживает макромолекулы. Истинный барьер, обеспечивающий избирательное поступление веществ в клетку, — ЦПМ. Она проницаема для одних веществ и непроницаема для других. Потоки веществ движутся в обоих направлениях (внутрь и наружу). Эти перемещения обеспечивают разнообразные транспортные системы, необходимые для выполнения двух важнейших задач. Обеспечение адекватных концентраций веществ, участвующих в основных биохимических реакциях, в том числе и обеспечение, при необходимости, их быстрого поступления внутрь клетки, невзирая на концентрацию этих веществ в окружающей среде. Поддержание осмотического давления, оптимального для протекания биохимических реакций. Поступление различных веществ внутрь бактериальной клетки реализуют три механизма: пассивный перенос, активный перенос и транспорт, обусловленный фосфорилированием Многие вещества способны неспецифически проникать в бактериальную клетку за счёт различия их концентраций по обе стороны ЦПМ. При этом они поступают в клетку только до выравнивания градиента концентрации с внешним раствором. Такое поступление веществ происходит пассивно, без прямых энергетических затрат. Существует два вида пассивной диффузии: простая и облегчённая. Простая диффузия. Проникновение веществ носит неспецифический характер и целиком зависит от размеров молекул и их липофильности. Скорость подобного переноса незначительна Транспорт сопровождается образованием комплекса «вещество-пермеаза». После преодоления ЦПМ комплекс диссоциирует, а пермеаза используется для последующего «проведения» других молекул. Подобный тип транспорта реализуется по градиенту концентрации и характерен для эукариотов при поглощении сахаров. У прокариотов единственный пример облегчённой диффузии — проникновение глицерина в клетки бактерий кишечной группы. При этом концентрация проникшего глицерина практически равна его концентрации в окружающей среде. В последующем (в результате реакций фосфорилирования) глицерин трансформируется в глицерин-3-фосфат. Активный перенос Концентрация некоторых веществ в бактериальной клетке может в сотни раз превышать их; содержание в окружающей среде. Транспорт таких веществ в клетку происходит против градиента концентрации перемещаемого вещества, требует затрат энергии и реализуется при помощи специфических переносчиков. Например, в случае активного транспорта, как и при облегчённой диффузии, перемещение конкретного вещества через ЦПМ осуществляет специфическая для такого вещества пермеаза. Активный транспорт сопровождается увеличением свободной энергии, которая составляет] 5, 71 lg C2/Cj кДж ¦ моль, где С2 и С( — соответственно более высокая и более низкая концен-; трации. Это обстоятельство делает процесс активного транспорта энергозависимым. У бактеА рий подобный тип поступления веществ доминирует, с его помощью транспортируются\ многие сахара, белки и другие вещества. Нередко у грамотрицательных бактерий в активном | переносе участвуют специальные белки, отличные от пермеаз, но действующие в ассоциации c l ними. Эти белки локализованы в периплазматическом пространстве. После проникновения вещества в клетку комплекс «субстрат — белок периплазмы — пермеаза» диссоциирует и для попавшего в клетку вещества вероятность выхода наружу резко снижается. Примером активного транспорта служит концентрирование лактозы в клетках кишечной палочки. Эта система известна как (З-галактозидпермеазная система. Энергия используется для снижения сродства пермеазы к лактозе на внутренней стороне ЦПМ по сравнению с ее сродством к тому же субстрату на внешней. В результате скорость выхода вещества наружу становится меньше, чем скорость его поступления внутрь клетки, и концентрация лактозы в клетке возрастает. Если блокировать образование энергии, то р-галактозидпермеазная система теряет способность осуществлять активный транспорт и переключается на облегчённую диффузию, обнаруживая одинаковое сродство к р-галактозидам по обе стороны мембраны. Транспорт, обусловленный фосфорилированием Транспорт, обусловленный фосфорилированием — энергозависимый процесс, используемый при утилизации углеводов. Основной механизм транспорта связан с фосфорилированием субстрата, что делает невозможным его выход из клетки. Первоначально происходит фосфори- лирование мембранного фермента (так называемый фермент 2, или Ф2) в цитоплазме за счёт фосфоенолпирувата. Фосфорилированный фермент связывает углевод (например, глюкозу или маннозу) на поверхности ЦПМ и транспортирует его в цитоплазму. Затем комплекс диссоциирует с высвобождением углевода, связанного с фосфатной группой. За счёт фосфорилирова- ния углевод аккумулируется в клетке и не способен выходить из неё. Данный тип транспорта не рассматривают как активный, поскольку концентрация неизменённого питательного вещества внутри клетки может быть одинаковой с его внеклеточным содержанием. Но в целом этот процесс напоминает активный транспорт против градиента концентрации вещества, так как концентрация химически изменённого питательного соединения внутри клетки может значительно превышать концентрацию неизменённого соединения в среде. Генетика бактерий Типы мутаций Термин «мутация» предложил де Фриз как понятие «скачкообразного изменения наследственного признака» при изучении наследственности у растений. Позднее Бёйеринк распространил это понятие и на бактерии. Мутация — изменение первичной структуры ДНК, проявляющееся наследственно закреплённой утратой или изменением какого-либо признака или группы признаков. Мутации подразделяют по происхождению, характеру изменений структуры ДНК, фенотипическим последствиям для клетки-мутанта и др. Факторы, вызывающие мутации, известны как мутагены. Они обычно имеют физическую или химическую природу. По происхождению выделяют мутации индуцированные, то есть вызванные искусственно, и спонтанные («дикие», возникают в популяции бактерий без видимого вмешательства извне). Спонтанные мутации К появлению спонтанных мутаций приводят ошибки репликации, неправильное формирование комплементарных пар оснований или структурные искажения ДНК под действием естественный мутагенов. Спонтанные мутации могут вызывать благоприятные и неблагоприятные генетические изменения. Примерный уровень спонтанного мутирования — одна мутация на каждые 106-107 клеток. Численная доля мутантов в клеточной популяции для разных признаков различна и может варьировать от 10" 4 до 10~п. Для конкретного гена частота мутирования составляет величину порядка 10“&, а для определённой пары нуклеотидов 10~8. Например, если на среду с антибиотиком посеять миллион бактерий, можно ожидать, что в результате спонтанной мутации одна колония выживет. Несмотря на то что уровень мутаций в популяции бактерий для отдельных клеток кажется незначительным, нужно помнить, что популяция бактерий огромна, и они размножается быстро. Следовательно, уровень мутаций с точки зрения целой популяции довольно значителен. Кроме того, появившиеся спонтанно и устойчивые к действию какого-либо антибиотика мутанты имеют при размножении преимущество по сравнению с «диким» типом бактерий и быстро образуют устойчивую популяцию. Обратные мутации {реверсии) возвращают спонтанно мутировавшую клетку к исходному генетическому состоянию. Их наблюдают с частотой одна клетка на 107—10е (то есть по меньшей мере в 10 раз реже, чем прямые спонтанные мутации). Индуцированные мутации Химический мутагенез. Некоторые химические вещества (мутагены) значительно повышают частоту мутирования до одной мутантной клетки на 103-104клеток. К таким веществам относят аналоги азотистых оснований (например, бромурацил), включающиеся в молекулу ДНК и вызывающие вставку некорректного основания при репликации (в частности, бромурацил аналогичен по структуре тимину, он включается в ДНК как партнёр аденина, а затем переходит в энольную форму и узнаётся полимеразой как цитозин, что приводит к включению гуанина вместо аденина); алкилирующие агенты (например, этилметансульфонат алкилиру- ет преимущественно атом азота гуанина); азотистая кислота, дезаминирующая азотистые основания; интеркалирующие агенты {например, акридиновые красители), внедряющиеся между основаниями ДНК и вызывающие увеличение расстояния между ними, что приводит к утрате нуклеотидов, включению дополнительной пары нуклеотидов и др. Радиационный мутагенез обычно приводит к образованию пиримидиновых димеров. УФ-, рентгеновские лучи и другие виды ионизирующего излучения оказывают на микроорганизмы как летальное (подавляющее жизнедеятельность), так и мутагенное воздействие. Мутации могут индуцировать следующие события: модификации оснований (изменения отдельных нуклеотидов), вставки (включение дополнительных оснований), делении (потеря одного основания или группы оснований) и деформации спирали ДНК. Модификация оснований включает химическое изменение азотистого основания в кодирующей последовательности, что приводит к изменению кодона. В результате вместо одной аминокислоты кодируется другая либо возникает бессмысленный кодон. Вставка либо делеция какого-либо основания (аналогов оснований) в ДНК приводит к фреймшифт-мутациям (мутации со сдвигом рамки считывания), что вызывает изменение позиции рамки считывания триплетного кодона, и, таким образом, изменение всех последующих кодонов. Деформации спирали ДНК (структурные искажения ДНК) образуются в результате индуцированной УФ-излучением димеризации расположенных близко нуклеотидов (особенно ти- мина). Образовавшееся циклобутановое кольцо нарушает симметрию ДНК и препятствует правильной репликации. Репликация может быть нарушена также при образовании поперечных межцепочечных сшивок ДНК. Б зависимости от синтеза «правильных» или «неправильных» полипептидов при считывании мРНК, отразившей изменения ДНК (то есть в зависимости от сохранения смысловой функции образующегося полипептида), различают несколько видов мутаций. «Молчащие» мутации (мутации «без изменения смысла», то есть не вызывающие изменения аминокислотной последовательности белка). Их появление возможно вследствие вырожденности генетического кода. Получившийся в результате мутирования триплет кодирует ту же самую аминокислоту, что и исходный триплет, поэтому синтезируемый белок остаётся без изменений. Миссенс-мутации (мутации «с изменением смысла») возникают при условии, что изменения кодирующей последовательности приводят к появлению в полипептиде иной аминокислоты. Получающийся изменённый белок может быть функциональным или нефункциональным в зависимости от значимости затронутой мутацией области. Нонсенс-мутации («антисмысловые», «бессмысленные» мутации) приводят к образованию одного из трёх кодонов-терминаторов (УАГ, УАА, УГА), вызывающих преждевременное окончание синтеза полипептидной цепи. Когда рибосома достигает такого кодона, процесс элонгации полипептидной цепи заканчивается, и высвобождается неполный пептид (вероятно, такое действие терминальных кодонов обусловлено отсутствием тРНК, связывающихся с данными кодонами). Эта мутация приводит либо к синтезу очень коротких нефункциональных белков, либо к полному прекращению синтеза белка.
ВЫВОДЫ Изучение морфолого-анатомических признаков бактерий помогает установить систематическое положение той или иной группы бактерий и, в конечном итоге определить их практическую значимость. По форме клеток бактерии разделяются: на шаровидные - кокки, палочковидные или цилиндрические - собственно бактерии и извитые - вибрионы спириллы и спирохеты. Между этими группами имеются переходные формы, например, кокко-бактерии. Бактерии являются прокариотами (рис. 1.1) и существенно отличаются от клеток растений и животных (эукариотов). Они относятся к одноклеточным организмам и состоят из клеточной стенки, цитоплазматической мембраны, цитоплазмы, нуклеоида (обязательных компонентов бактериальной клетки). Некоторые бактерии могут иметь жгутики, капсулы, споры (необязательные компоненты бактериальной клетки). Движение бактерий не является особенностью всех бактерий, только некоторые способны двигаться. Процесс спорообразования у бактерий является стадией развития. Спора играет роль защитного приспособления, которым природа снабдила их для борьбы с неблагоприятными условиями. Способностью образовывать споры - особые тельца эллиптической и округлой формы обладают палочковидные бактерии. Основным путем размножения у бактерий является бесполое размножение. Однако у некоторых бактерий встречается и половое размножение - конъюгация. Жизнь бактерий, как и других живых существ, в упрощённом виде сводится к таким последовательным событиям: собственное воспроизведение -» обеспечение жизненных функций -» воспроизведение потомства. Нормальная реализация этого круговорота жизни для любого вида бактерий возможна лишь при развитии адекватных ответных реакций на любые воздействия окружающей среды, что во многом обеспечивается генетической информацией организма. Жизнь бактерий, равно как и всех живых существ, характеризуется ростом и размножением, то есть увеличением живой массы отдельной особи и популяции в целом за счет ассимиляции веществ, находящихся вне клетки. Бактерии — удобная модель для генетических исследований.
СПИСОК ЛИТЕРАТУРЫ 1.Морфология и анатомия высших растений. Рубрика: Естественные науки. Название: Морфология и анатомия высших растений. Автор: А. А. Седов 2.Медицинская микробиология: конспект лекций для вузов В.А. Подколзина, 3. Микробиология - Дикий И.Л. 4. Руководство к лабораторным занятиям «Микробиология» Лысак В.Ю. 5. Ксения Викторовна Ткаченко «Микробиология» 6. Интернет сайт http: //bibliofond.ru/view.aspx? id=606606 7. Интернет сайт http: //www.studfiles.ru/preview/1777984/ 8. Интернет сайт http: //medbe.ru/materials/mikrobiologiya-i-biotekhnologii/ 9. Интернет сайт http: //atlas.ztom.ru/index.php/morfologiya-mm/anatomiya-bak-kletki-mm 10. Интернет сайт http: //studopedia.ru/3_25993_morfologiya-i-ultrastruktura-bakterialnih-kletok.html
ВВЕДЕНИЕ Изучение морфолого-анатомических признаков бактерий помогает установить систематическое положение той или иной группы бактерий и, в конечном итоге определить их практическую значимость. Небольшие размеры бактерий долгое время казались несовместимыми с какой-либо клеточной дифференцировкой. Использование новейших методов исследования позволило выделить клеточные структуры изучить их физиологические функции. Большую роль в изучении бактерий сыграли косвенные методы, основанные на способности клетки реагировать на иммунные антитела. Оказалось, что реагируют в клетке белки с определенной формой и расположением полипептидной цепи. Это позволило изучить то, что лежит за пределами видимости микроскопа. Популярное:
|
Последнее изменение этой страницы: 2016-06-05; Просмотров: 884; Нарушение авторского права страницы